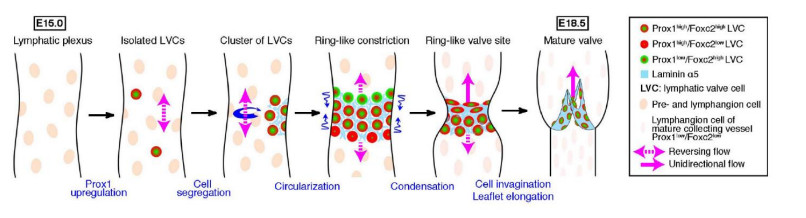
A majority of lymphatic valves tend to form in proximity to vessel junctions, and it is often proposed that disturbed flow at junctions creates oscillating shear stress that leads to accumulation of transcription factors which bring about valvogenesis at these sites. In images of networks of dorsal skin lymphatics from embryonic mice (day E16), we compared simulated fluid flow patterns and observed distributions of the transcription factor Prox1, which is implicated in valve formation. Because of creeping-flow conditions, flow across vessel junctions was not 'disturbed', and within a given vessel, shear stress varied inversely with local conduit width. Prox1 concentration was indeed localised to vessel end-regions, but over three networks was not consistently correlated with the vessel normalised-distance distribution of either fluid shear stress or shear-stress axial gradient. These findings do not support the presently accepted mechanism for the role of flow in valve localisation.
Citation: Christopher D. Bertram, Bernard O. Ikhimwin, Charlie Macaskill. Modeling flow in embryonic lymphatic vasculature: what is its role in valve development?[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1406-1424. doi: 10.3934/mbe.2021073
A majority of lymphatic valves tend to form in proximity to vessel junctions, and it is often proposed that disturbed flow at junctions creates oscillating shear stress that leads to accumulation of transcription factors which bring about valvogenesis at these sites. In images of networks of dorsal skin lymphatics from embryonic mice (day E16), we compared simulated fluid flow patterns and observed distributions of the transcription factor Prox1, which is implicated in valve formation. Because of creeping-flow conditions, flow across vessel junctions was not 'disturbed', and within a given vessel, shear stress varied inversely with local conduit width. Prox1 concentration was indeed localised to vessel end-regions, but over three networks was not consistently correlated with the vessel normalised-distance distribution of either fluid shear stress or shear-stress axial gradient. These findings do not support the presently accepted mechanism for the role of flow in valve localisation.

| [1] |
A. Sabine, Y. Agalarov, H. M-E. Hajjami, M. Jaquet, R. Hägerling, C. Pollmann, et al., Mechanotransduction, PROX1, and FOXC2 cooperate to control connexin37 and calcineurin during lymphatic-valve formation, Dev. Cell, 22 (2012), 430-445. doi: 10.1016/j.devcel.2011.12.020

|
| [2] |
L. Planas-Paz, E. Lammert, Mechanical forces in lymphatic vascular development and disease, Cell. Mol. Life Sci., 70 (2013), 4341-4354. doi: 10.1007/s00018-013-1358-5
|
| [3] |
D. Choi, E. Park, E. Jung, B. Cha, S. Lee, J. Yu, et al., Piezo1 incorporates mechanical force signals into the genetic program that governs lymphatic valve development and maintenance, JCI Insight, 4 (2019), e125068-1-e125068-15. doi: 10.1172/jci.insight.125068
|
| [4] |
O. F. Kampmeier, The genetic history of the valves in the lymphatic system of man. Am. J. Anat., 40 (1928), 413-457. doi: 10.1002/aja.1000400302
|
| [5] |
E. Bazigou, J. T. Wilson, J. E. Moore jr., Primary and secondary lymphatic valve development: molecular, functional and mechanical insights, Microvasc. Res., 96 (2014), 38-45. doi: 10.1016/j.mvr.2014.07.008
|
| [6] |
D. T. Sweet, J. M. Jiménez, J. Chang, P. R. Hess, P. Mericko-Ishizuka, J. Fu, et al., Lymph flow regulates collecting lymphatic vessel maturation in vivo, J. Clin. Invest., 125 (2015), 2995-3007. doi: 10.1172/JCI79386
|
| [7] |
B. K. Bharadvaj, R. F. Mabon, D. P. Giddens, Steady flow in a model of the human carotid bifurcation. Part I——Flow visualization, J. Biomech., 15 (1982), 349-362. doi: 10.1016/0021-9290(82)90057-4
|
| [8] |
B. K. Bharadvaj, R. F. Mabon, D. P. Giddens, Steady flow in a model of the human carotid bifurcation. Part Ⅱ——Laser Doppler anemometer measurements, J. Biomech., 15 (1982), 363-378. doi: 10.1016/0021-9290(82)90058-6
|
| [9] |
D. N. Ku, D. P. Giddens, Pulsatile flow in a model carotid bifurcation, Arteriosclerosis, 3 (1983), 31-39. doi: 10.1161/01.ATV.3.1.31
|
| [10] |
D. N. Ku, D. P. Giddens, Laser Doppler anemometer measurements of pulsatile flow in a model carotid bifurcation, J. Biomech., 20 (1987), 407-421. doi: 10.1016/0021-9290(87)90048-0
|
| [11] |
D. N. Ku, D. P. Giddens, D. J. Phillips, D. E. Strandness jr., Hemodynamics of the normal human carotid bifurcation: in vitro and in vivo studies, Ultrasound Med. Biol., 11 (1985), 13-26. doi: 10.1016/0301-5629(85)90003-1
|
| [12] |
D. N. Ku, D. P. Giddens, C. K. Zarins, S. Glagov, Pulsatile flow and atherosclerosis in the human carotid bifurcation, Arteriosclerosis, 5 (1985), 293-302. doi: 10.1161/01.ATV.5.3.293
|
| [13] |
C. Hahn, M. A. Schwartz, Mechanotransduction in vascular physiology and atherogenesis, Nat. Rev. Mol. Cell Bio., 10 (2009), 53-62. doi: 10.1038/nrm2596
|
| [14] |
J. Kazenwadel, K. L. Betterman, C. E. Chong, P. H. Stokes, Y. K. Lee, G. A. Secker, et al., GATA2 is required for lymphatic vessel valve development and maintenance, J. Clin. Invest., 125 (2015), 2979-2994. doi: 10.1172/JCI78888
|
| [15] |
P. R. Norden, A. Sabine, Y. Wang, C. S. Demir, T. Liu, T. V. Petrova, et al., Shear stimulation of FOXC1 and FOXC2 differentially regulates cytoskeletal activity during lymphatic valve maturation, eLife, 9 (2020), e53814-1-e53814-35. doi: 10.7554/eLife.53814
|
| [16] |
C. Norrmén, K. I. Ivanov, J. Cheng, N. Zangger, M. Delorenzi, M. Jaquet, et al., FOXC2 controls formation and maturation of lymphatic collecting vessels through cooperation with NFATc1, J. Cell Biol., 185 (2009), 439-457. doi: 10.1083/jcb.200901104
|
| [17] |
J. E. Moore jr., C. D. Bertram, Lymphatic system flows, Annu. Rev. Fluid Mech., 50 (2018), 459-482. doi: 10.1146/annurev-fluid-122316-045259
|
| [18] | A. Sabine, T. V. Petrova, Interplay of mechanotransduction, FOXC2, connexins, and calcineurin signaling in lymphatic valve formation. In: Developmental Aspects of the Lymphatic Vascular System (eds. F Kiefer, S Schulte-Merker) Advances in Anatomy, Embryology and Cell Biology, vol 214. Springer-Verlag, Wien, (2014), pp 67-80.doi:10.1007/978-3-7091-1646-3_6 |
| [19] |
J. M. Dolan, H. Meng, S. Singh, R. Paluch, J. Kolega, High fluid shear stress and spatial shear stress gradients affect endothelial proliferation, survival, and alignment, Ann. Biomed. Eng., 39 (2011), 1620-1631. doi: 10.1007/s10439-011-0267-8
|
| [20] |
J. M. Dolan, H. Meng, F. J. Sim, J. Kolega, Differential gene expression by endothelial cells under positive and negative streamwise gradients of high wall shear stress, Am. J. Physiol. Cell Physiol., 305 (2013), C854-C866. doi: 10.1152/ajpcell.00315.2012
|
| [21] |
J. M. Dolan, J. Kolega, H. Meng, High wall shear stress and spatial gradients in vascular pathology: a review, Ann. Biomed. Eng., 41 (2013), 1411-1427. doi: 10.1007/s10439-012-0695-0
|
| [22] |
V. N. Surya, E. Michalaki, G. G. Fuller, A. R. Dunn, Lymphatic endothelial cell calcium pulses are sensitive to spatial gradients in wall shear stress, Mol. Biol. Cell, 30 (2019), 923-931. doi: 10.1091/mbc.E18-10-0618
|
| [23] |
E. Michalaki, V. N. Surya, G. G. Fuller, A. R. Dunn, Perpendicular alignment of lymphatic endothelial cells in response to spatial gradients in wall shear stress, Commun. Biol., 3 (2020), 57-1-57-9. doi: 10.1038/s42003-019-0732-8
|
| [24] | A. Sabine, M. J. Davis, E. Bovay, T. V. Petrova, Characterization of mouse mesenteric lymphatic valve structure and function. In: Lymphangiogenesis (eds. G Oliver, M Kahn). Methods in Molecular Biology, vol 1846. Humana Press, New York, (2018), pp 97-129.doi:10.1007/978-1-4939-8712-2_7 |
| [25] |
M. R. Kelly-Goss, E. R. Winterer, P. C. Stapor, M. Yang, R. S. Sweat, W. B. Stallcup, et al., Cell proliferation along vascular islands during microvascular network growth, BMC Physiol., 12 (2012), 7-1-7-9. doi: 10.1186/1472-6793-12-7
|
| [26] |
J. Chiu, S. Chien, Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives, Physiol. Rev., 91 (2011), 327-387. doi: 10.1152/physrev.00047.2009
|














Figures(15) / Tables(1)

Christopher D. Bertram, Bernard O. Ikhimwin, Charlie Macaskill. Modeling flow in embryonic lymphatic vasculature: what is its role in valve development?[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1406-1424. doi: 10.3934/mbe.2021073







 DownLoad:
DownLoad: