In this study, we explore a stochastic age-dependent cooperative Lotka-Volterra (LV) system with an environmental noise. By applying the theory of M-matrix, we prove the existence and uniqueness of the global solution for the system. Since the stochastic age-dependent cooperative LV system cannot be solved explicitly, we then construct an Euler-Maruyama (EM) numerical solution to approach the exact solution of the system. The convergence rate and the $ p $th-moment boundedness of the scheme have also been obtained. Additionally, numerical experiments have been conducted to verify our theoretical results.
Citation: Mengqing Zhang, Qimin Zhang, Jing Tian, Xining Li. The asymptotic stability of numerical analysis for stochastic age-dependent cooperative Lotka-Volterra system[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1425-1449. doi: 10.3934/mbe.2021074
In this study, we explore a stochastic age-dependent cooperative Lotka-Volterra (LV) system with an environmental noise. By applying the theory of M-matrix, we prove the existence and uniqueness of the global solution for the system. Since the stochastic age-dependent cooperative LV system cannot be solved explicitly, we then construct an Euler-Maruyama (EM) numerical solution to approach the exact solution of the system. The convergence rate and the $ p $th-moment boundedness of the scheme have also been obtained. Additionally, numerical experiments have been conducted to verify our theoretical results.

| [1] |
J. H. Vandermeer, D. H. Boucher, Varieties of mutualistic interaction in population models, J. Theor. Biol., 74 (1978), 549–558. doi: 10.1016/0022-5193(78)90241-2

|
| [2] |
W. J. Zuo, D. Q. Jiang, X. G. Sun, T. Hayat, A. Alsaedi, Long-time behaviors of a stochastic cooperative Lotka-Volterra system with distributed delay, Physica A., 506 (2018), 542–559. doi: 10.1016/j.physa.2018.03.071
|
| [3] | Q. Liu, Analysis of a stochastic non-autonomous food-limited Lotka-Volterra cooperative model, Appl. Math. Comput., 254 (2015), 1–8. |
| [4] | E. P. Odum, Fundamentals of Ecology 5th Edition, Philadelphia, Saunders, 1971. |
| [5] | J. D. Murray, Mathematical Biology, Springer, Berlin, 1989. |
| [6] | M. E. Hale, The Biology of Lichens, Arnold, London, 1974. |
| [7] |
J. Roughgarden, Evolution of marine symbiosis-A simple cost-benefit model, Ecology, 56 (1975), 1201–1208. doi: 10.2307/1936160
|
| [8] |
J. H. Huang, X. F. Zou, Traveling wavefronts in diffusive and cooperative Lotka-Volterra system with delays, J. Math. Anal. Appl., 271 (2002), 455–466. doi: 10.1016/S0022-247X(02)00135-X
|
| [9] |
G. C. Lu, Z. Y. Lu, Permanence for two-species Lotka-Volterra cooperative systems with delays, Math. Biosci. Eng., 5 (2008), 477–484. doi: 10.3934/mbe.2008.5.477
|
| [10] | G. C. Lu, Z. Y. Lu, X. Z. Lian, Delay effect on the permanence for Lotka-Volterra cooperative systems, Nonlinear. Anal-Real., 11 (2010), 2810–2816. |
| [11] | Y. Nakata, Y. Muroya, Permanence for nonautonomous Lotka-Volterra cooperative systems with delays, Nonlinear. Anal-Real., 11 (2010), 528–534. |
| [12] | L. Xu, J. Y. Liu, G. Zhang, Pattern formation and parameter inversion for a discrete Lotka-Volterra cooperative system, Chaos. Soliton. Fract., 110 (2018), 226–231. |
| [13] | W. T. Li, Z. C. Wang, Traveling fronts in diffusive and cooperative Lotka-Volterra system with nonlocal delays, Z. Angew. Math. Phys., 58 (2007), 571–591. |
| [14] | A. J. Lotka, Relation between birth rates and death rates, Science, 26 (1907), 121–130. |
| [15] | X. R. Mao, Stochastic Differential Equations and Applications, Horwood, UK, 2007. |
| [16] | X. M. Zhang, Z. H. Liu, Periodic oscillations in age-structured ratio-dependent predator-prey model with Michaelis-Menten type functional response, Physica D., 389 (2019), 51–63. |
| [17] | F. J. Solis, R. A. Ku-Carrillo, Generic predation in age structure predator-prey models, Appl. Math. Comput., 231 (2014), 205–213. |
| [18] |
M. Delgado, A. Suárez, Age-dependent diffusive Lotka-Volterra type systems, Math. Comput. Model, 45 (2007), 668–680. doi: 10.1016/j.mcm.2006.07.013
|
| [19] | N. Hritonenko, Y. Yatsenko, The structure of optimal time and age-dependent harvesting in the Lotka-McKendrik population model, Math. Biosci., 208 (2007), 48–62. |
| [20] | D. W. Tudor, An age-dependent epidemic model with application to measles, Math. Biosci., 73 (1985), 131–147. |
| [21] | D. Greenhalgh, Threshold and stability results for an epidemic model with an age-structured meeting rate, Math. Med. Biol., 5 (1988), 81–100. |
| [22] | Q. M. Zhang, W. A. Liu, Z. K. Nie, Existence, uniqueness and exponential stability for stochastic age-dependent population, Appl. Math. Comput., 154 (2004), 183–201. |
| [23] | Y. Li, M. Ye, Q. M. Zhang, Strong convergence of the partially truncated Euler-Maruyama scheme for a stochastic age-structured SIR epidemic model, Appl. Math. Comput., 362 (2019), 1–22. |
| [24] | W. K. Pang, R. H. Li, M. Liu, Exponential stability of numerical solutions to stochastic age-dependent population equations, Appl. Math. Comput., 183 (2006), 152–159. |
| [25] | R. H. Li, H. B. Meng, Q. Chang, Convergence of numerical solutions to stochastic age-dependent population equations, J. Comput. Appl. Math., 193 (2006), 109–120. |
| [26] | R. H. Li, P. K. Leung, W. K. Pang, Convergence of numerical solutions to stochastic age-dependent population equations with Markovian switching, J. Comput. Appl. Math., 233 (2009), 1046–1055. |
| [27] | R. H. Li, W. K. Pang, Q. H. Wang, Numerical analysis for stochastic age-dependent population equations with Poisson jumps, J. Math. Anal. Appl., 327 (2007), 1214–1224. |
| [28] | W. K. Pang, R. H. Li, M. Liu, Convergence of the semi-implicit Euler method for stochastic age-dependent population equations, Appl. Math. Comput., 195 (2008), 466–474. |
| [29] | X. Y. Li, Variational iteration method for nonlinear age-structured population models, Comput. Math. Appl., 58 (2009), 2177–2181. |
| [30] | Y. Yang, C. F. Wu, Z. X. Li, Forced waves and their asymptotics in a Lotka-Volterra cooperative model under climate change, Appl. Math. Comput., 353 (2019), 254–264. |
| [31] | Y. Zhao, S. L. Yuan, Q. M. Zhang, The effect of noise on the survival of a stochastic competitive model in an impulsive polluted environment, Appl. Math. Model., 40 (2016), 7583–7600. |
| [32] | C. Q. Xu, S. L. Yuan, T. H. Zhang, Global dynamics of a predator-prey model with defence mechanism for prey, Appl. Math. Lett., 62 (2016), 42–48. |
| [33] | Q. M. Zhang, Exponential stability of numerical solutions to a stochastic age-structured population system with diffusion, J. Comput. Appl. Math., 220 (2008), 22–33. |
| [34] |
A. Chekroun, T. Kuniya, Global threshold dynamics of an infection age-structured SIR epidemic model with diffusion under the Dirichlet boundary condition, J. Diff. Equations, 269 (2020), 117–148. doi: 10.1016/j.jde.2020.04.046
|
| [35] | R. X. Lu, F. Y. Wei, Persistence and extinction for an age-structured stochastic SVIR epidemic model with generalized nonlinear incidence rate, Physica A., 513 (2019), 572–587. |
| [36] | P. Yang, Y. S. Wang, Existence and properties of Hopf bifurcation in an age-dependent predation system with prey harvesting, Commun. Nonlinear. Sci. Numer. Simulat., 91 (2020), 105395. |
| [37] | Z. X. Luo, Z. R. He, Optimal control for age-dependent population hybrid system in a polluted environment, Appl. Math. Comput., 228 (2014), 68–76. |
| [38] | C. Burgos, J. C. Cortés, L. Shaikhet, R. J. Villanueva, A nonlinear dynamic age-structured model of e-commerce in spain: Stability analysis of the equilibrium by delay and stochastic perturbations, Commun. Nonlinear. Sci. Numer. Simulat., 64 (2018), 149–158. |
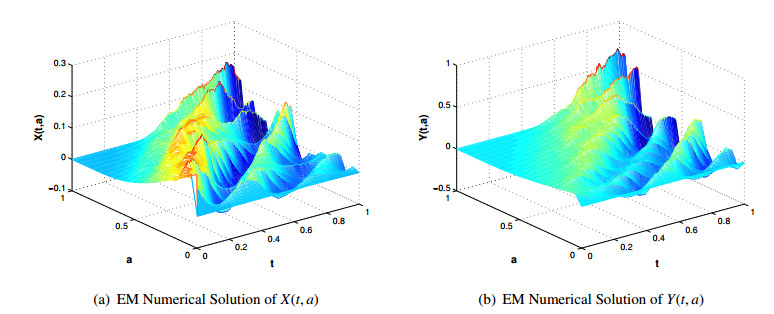
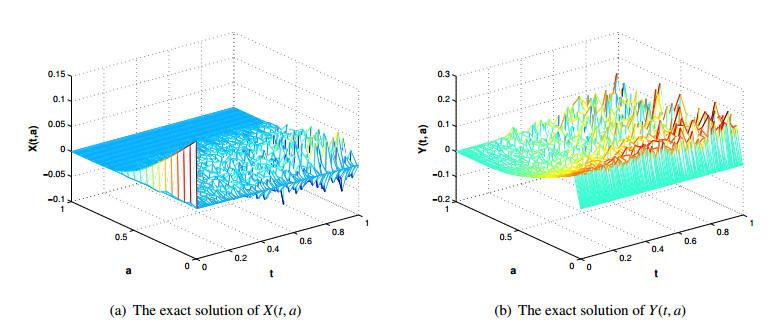
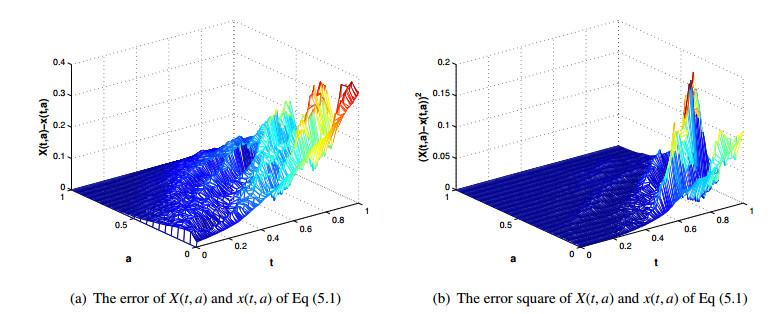
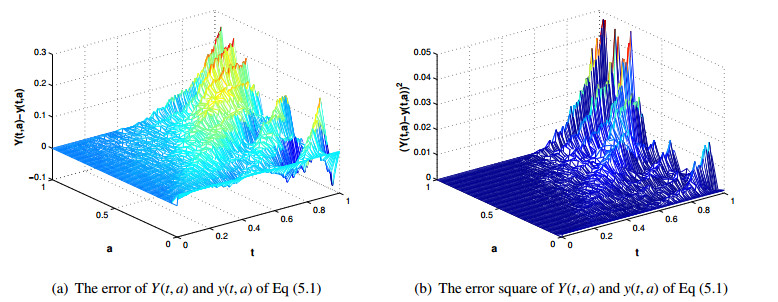
Figures(5) / Tables(3)

Mengqing Zhang, Qimin Zhang, Jing Tian, Xining Li. The asymptotic stability of numerical analysis for stochastic age-dependent cooperative Lotka-Volterra system[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1425-1449. doi: 10.3934/mbe.2021074







 DownLoad:
DownLoad: