Citation: Tomás Caraballo, Renato Colucci, Javier López-de-la-Cruz, Alain Rapaport. Study of the chemostat model with non-monotonic growth under random disturbances on the removal rate[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7480-7501. doi: 10.3934/mbe.2020382

| [1] | J. Monod, La technique de culture continue: Théorie et applications, Annales de l'Institute Pasteur, 79 (1950), 390-410. |
| [2] |
A. Novick, L. Szilard, Experiments with the chemostat on spontaneous mutations of bacteria, Proc. Natl. Acad. Sci., 36 (1950), 708-719. doi: 10.1073/pnas.36.12.708

|
| [3] |
H. W. Jannasch, Steady state and the chemostat in ecology, Limnol. Oceanogr., 19 (1974), 716- 720. doi: 10.4319/lo.1974.19.4.0716
|
| [4] |
G. D'Ans, P. Kokotovic, D. Gottlieb, A nonlinear regulator problem for a model of biological waste treatment, IEEE Trans. Autom. Control., 16 (1971), 341-347. doi: 10.1109/TAC.1971.1099745
|
| [5] | J. W. M. La Rivière, Microbial ecology of liquid waste treatment, in Advances in Microbial Ecology, 1 (1977), 215-259, Springer US. |
| [6] | J. Barlow, F. de Noyelles, B. Peterson, J. Peterson, W. Schaffner, "Continuous flow nutrient bioassays with natural phytoplankton populations". G. Glass (Editor): Bioassay Techniques and Environmental Chemistry, John Wiley & Sons Ltd., 1973. |
| [7] |
I. F. Creed, D. M. McKnight, B. A. Pellerin, M. B. Green, B. A. Bergamaschi, G. R. Aiken, et al., The river as a chemostat: fresh perspectives on dissolved organic matter flowing down the river continuum, Can. J. Fish. Aquatic Sci., 72 (2015), 1272-1285. doi: 10.1139/cjfas-2014-0400
|
| [8] | S. Jorgensen, B. Fath, Fundamentals of Ecological Modelling Applications in Environmental Management and Research. Elsevier, 2011. |
| [9] |
J. Kalff, R. Knoechel, Phytoplankton and their dynamics in oligotrophic and eutrophic lakes, Annu. Rev. Ecol. Syst., 9 (1978), 475-495. doi: 10.1146/annurev.es.09.110178.002355
|
| [10] |
E. Rurangwa, M. C. J. Verdegem, Microorganisms in recirculating aquaculture systems and their management, Rev. Aquac., 7 (2015), 117-130. doi: 10.1111/raq.12057
|
| [11] | J. Harmand, C. Lobry, A. Rapaport, T. Sari, The Chemostat: Mathematical Theory of Microorganisms Cultures. Wiley, Chemical Engineering Series, John Wiley & Sons, Inc., 2017. |
| [12] | H. L. Smith, P. Waltman, The theory of the chemostat: dynamics of microbial competition. Cambridge University Press, 1995. |
| [13] | T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz, A. Rapaport, Modeling and analysis of random and stochastic input flows in the chemostat model, Discret. Continuous Dyn. Syst. Ser. B, 24 (2018), 3591-3614. |
| [14] | Y. Asai, P. Kloeden, Numerical schemes for random odes via stochastic differential equations, Commun. Appl. Analysis, 17 (2013), 521-528. |
| [15] |
T. Caraballo, X. Han, P. E. Kloeden, Chemostats with random inputs and wall growth, Math. Methods Appl. Sci., 38 (2015), 3538-3550. doi: 10.1002/mma.3437
|
| [16] | T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz, Some Aspects Concerning the Dynamics of Stochastic Chemostats, vol. 69, ch. 11, pp. 227-246. Springer International Publishing, 2016. |
| [17] |
T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz, Dynamics of some stochastic chemostat models with multiplicative noise, Commun. Pure Appl. Analysis, 16 (2017), 1893-1914. doi: 10.3934/cpaa.2017092
|
| [18] |
J. Andrews, A mathematical model for the continuous culture of microorganisms utilizing inhibitory substrates, Biotechnol. Bioeng., 10 (1968), 707-723. doi: 10.1002/bit.260100602
|
| [19] | G. Bastin, D. Dochain, On-line estimation and adaptive control of bioreactors. Elsevier, 1990. |
| [20] |
A. Rapaport, J. Harmand, Robust regulation of a class of partially observed nonlinear continuous bioreactors, J. Process. Control., 12 (2002), 291-302. doi: 10.1016/S0959-1524(01)00029-4
|
| [21] |
B. Satishkumar, M. Chidambaram, Control of unstable bioreactor using fuzzy tuned PI controller, Bioprocess Eng., 20, (1999), 127-132. doi: 10.1007/s004490050570
|
| [22] |
A. Schaum, J. Alvarez, T. Lopez-Arenas, Saturated PI control of continuous bioreactors with haldane kinetics, Chem. Eng. Sci., 68 (2012), 520-529. doi: 10.1016/j.ces.2011.10.006
|
| [23] | A. Rapaport, I. Haidar, J. Harmand, Global dynamics of the buffered chemostat for a general class of response functions, J. Math. Biol., 71 (2014), 69-98. |
| [24] |
A. Rapaport, J. Harmand, Biological control of the chemostat with nonmonotonic response and different removal rates, Math. Biosci. Eng., 5 (2008), 539-547. doi: 10.3934/mbe.2008.5.539
|
| [25] |
B. Cloez, C. Fritsch, Gaussian approximations for chemostat models in finite and infinite dimensions, J. Math. Biol., 75 (2017), 805-843. doi: 10.1007/s00285-017-1097-6
|
| [26] |
P. Collet, S. Martínez, S. Méléard, J. S. Martín, Stochastic models for a chemostat and long-time behavior, Adv. Appl. Probab., 45 (2013), 822-836. doi: 10.1017/S0001867800006595
|
| [27] |
C. Fritsch, J. Harmand, F. Campillo, A modeling approach of the chemostat, Ecol. Model., 299 (2015), 1-13. doi: 10.1016/j.ecolmodel.2014.11.021
|
| [28] |
J. Grasman, M. D. Gee, O. A. V. Herwaarden, Breakdown of a chemostat exposed to stochastic noise, J. Eng. Math., 53 (2005), 291-300. doi: 10.1007/s10665-005-9004-3
|
| [29] |
L. Imhof, S. Walcher, Exclusion and persistence in deterministic and stochastic chemostat models, J. Differ. Equations, 217 (2005), 26-53. doi: 10.1016/j.jde.2005.06.017
|
| [30] |
G. Stephanopoulos, R. Aris, A. Fredrickson, A stochastic analysis of the growth of competing microbial populations in a continuous biochemical reactor, Math. Biosci., 45 (1979), 99-135. doi: 10.1016/0025-5564(79)90098-1
|
| [31] |
C. Xu, S. Yuan, An analogue of break-even concentration in a simple stochastic chemostat model, Appl. Math. Lett., 48 (2015), 62-68. doi: 10.1016/j.aml.2015.03.012
|
| [32] |
D. Zhao, S. Yuan, Critical result on the break-even concentration in a single-species stochastic chemostat model, J. Math. Anal. Appl., 434 (2016), 1336-1345. doi: 10.1016/j.jmaa.2015.09.070
|
| [33] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz, A. Rapaport, A way to model stochastic perturba-tions in population dynamics models with bounded realizations, Commun. Nonlinear Sci. Numer. Simul., 77 (2019), 239-257. doi: 10.1016/j.cnsns.2019.04.019
|
| [34] | T. Caraballo, X. Han, Applied Nonautonomous and Random Dynamical Systems, Applied Dynamical Systems. Springer International Publishing, 2016. |
| [35] | L. Arnold, Random Dynamical Systems. Springer Berlin Heidelberg, 1998. |
| [36] |
T. Caraballo, P. E. Kloeden, B. Schmalfuss, Exponentially stable stationary solutions for stochastic evolution equations and their perturbation, Appl. Math. Optim., 50 (2004), 183-207. doi: 10.1007/s00245-004-0802-1
|
| [37] |
H. I. Freedman, P. Moson, Persistence definitions and their connections, Proc. Am. Math. Soc., 109 (1990), 1025-1033. doi: 10.1090/S0002-9939-1990-1012928-6
|
| [38] | W. Walter, Ordinary Differential Equations. Springer New York, 1998. |
| [39] | H. R. Thieme, Convergence results and a poincare-bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755-763. |
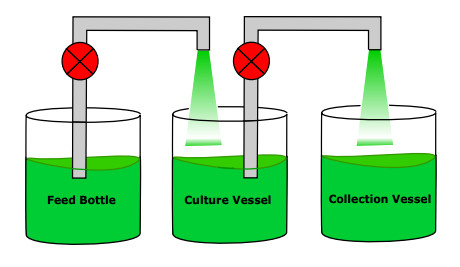
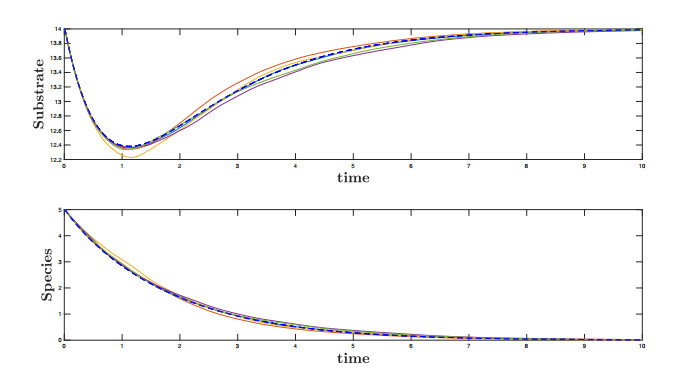
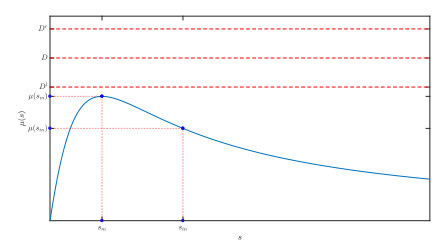
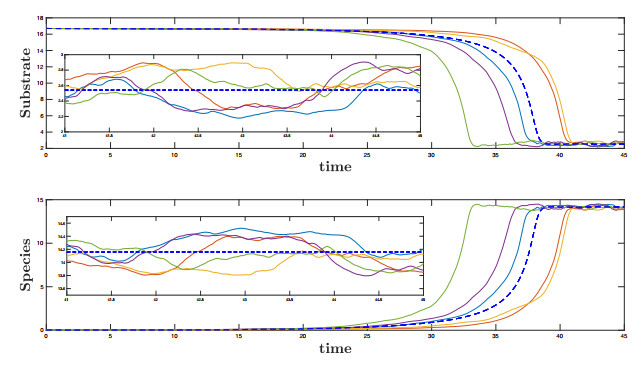
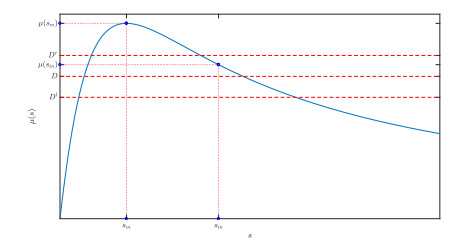
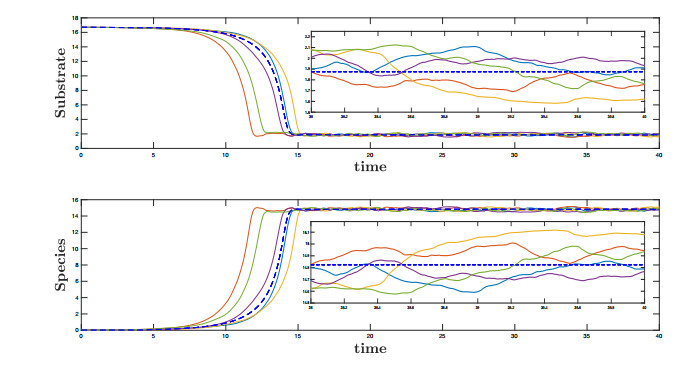
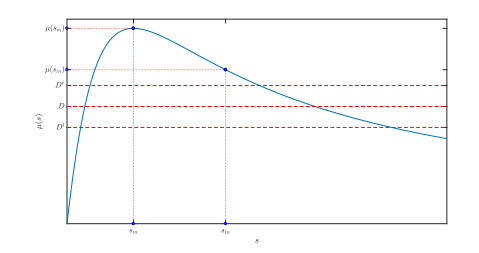
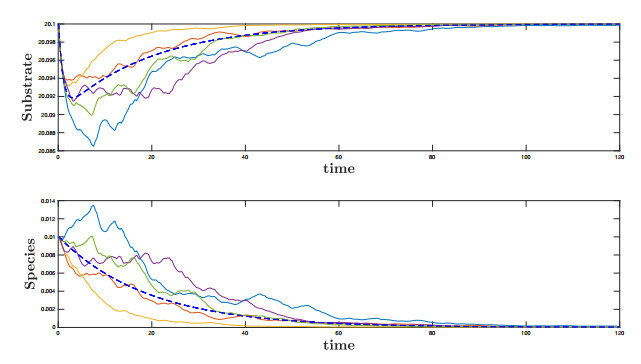
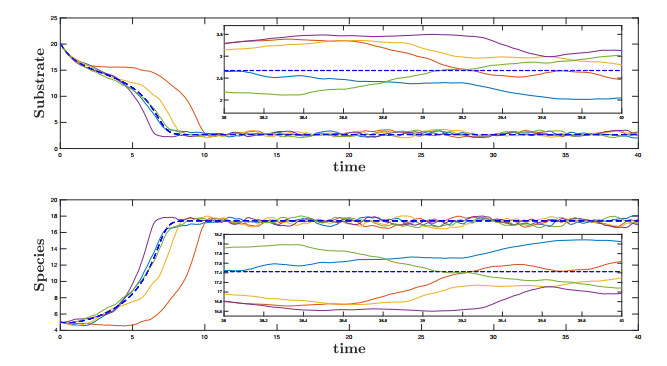
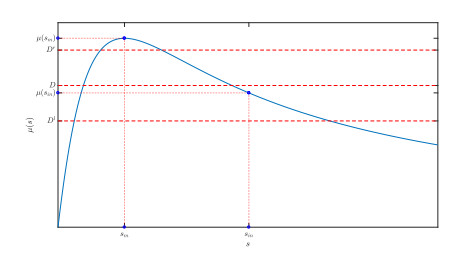
Figures(10)

Tomás Caraballo, Renato Colucci, Javier López-de-la-Cruz, Alain Rapaport. Study of the chemostat model with non-monotonic growth under random disturbances on the removal rate[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7480-7501. doi: 10.3934/mbe.2020382







 DownLoad:
DownLoad: