Citation: Fiona R. Macfarlane, Mark A. J. Chaplain, Tommaso Lorenzi. A hybrid discrete-continuum approach to model Turing pattern formation[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7442-7479. doi: 10.3934/mbe.2020381

| [1] |
A. M. Turing, The chemical basis of morphogenesis, Philos. Trans. R. Soc. Lond. B, 237 (1952), 37-72. doi: 10.1098/rstb.1952.0012

|
| [2] |
A. Gierer and H. Meinhardt, A theory of biological pattern formation, Kybernetik, 12 (1972), 30-39. doi: 10.1007/BF00289234
|
| [3] | H. Meinhardt, Models of Biological Pattern Formation, Academic Press, London, 1982. |
| [4] |
J. D. Murray, A pre-pattern formation mechanism for animal coat markings, J. Theor. Biol., 88 (1981), 161-199. doi: 10.1016/0022-5193(81)90334-9
|
| [5] | P. K. Maini and T. E. Woolley, The Turing model for biological pattern formation, in The Dynamics of Biological Systems, Springer, 2019,189-204. |
| [6] | P. Arcuri and J. D. Murray, Pattern sensitivity to boundary and initial conditions in reaction-diffusion models, J. Math. Biol., 24 (1986), 141-165. |
| [7] |
M. A. J. Chaplain, M. Ganesh, and I. G. Graham, Spatio-temporal pattern formation on spherical surfaces: Numerical simulation and application to solid tumour growth, J. Math. Biol., 42 (2001), 387-423. doi: 10.1007/s002850000067
|
| [8] |
E. J. Crampin, E. A. Gaffney, and P. K. Maini, Reaction and diffusion on growing domains: Scenarios for robust pattern formation, Bull. Math. Biol., 61 (1999), 1093-1120. doi: 10.1006/bulm.1999.0131
|
| [9] |
E. J. Crampin, W. W. Hackborn, and P. K. Maini, Pattern formation in reaction-diffusion models with nonuniform domain growth, Bull. Math. Biol., 64 (2002), 747-769. doi: 10.1006/bulm.2002.0295
|
| [10] |
S. Kondo and R. Asai, A reaction-diffusion wave on the skin of the marine angelfish Pomacanthus, Nature, 376 (1995), 765-768. doi: 10.1038/376765a0
|
| [11] |
A. L. Krause, M. A. Ellis, and R. A. Van Gorder. Influence of curvature, growth, and anisotropy on the evolution of Turing patterns on growing manifolds, Bull. Math. Biol., 81 (2019), 759-799. doi: 10.1007/s11538-018-0535-y
|
| [12] |
A. L. Krause, V. Klika, T. E. Woolley, and E. A. Gaffney, From one pattern into another: Analysis of Turing patterns in heterogeneous domains via WKBJ, J. R. Soc. Interface, 17 (2020), 20190621. doi: 10.1098/rsif.2019.0621
|
| [13] |
P. K. Maini, T. E. Woolley, R. E. Baker, E. A. Gaffney, and S. S. Lee, Turing's model for biological pattern formation and the robustness problem, Interface Focus, 2 (2012), 487-496. doi: 10.1098/rsfs.2011.0113
|
| [14] |
A. Madzvamuse, A. H. W. Chung, and C. Venkataraman, Stability analysis and simulations of coupled bulk-surface reaction-diffusion systems, Proc. R. Soc. A, 471 (2015), 20140546. doi: 10.1098/rspa.2014.0546
|
| [15] |
A. Madzvamuse, A. J. Wathen, and P. K. Maini, A moving grid finite element method applied to a model biological pattern generator, J. Comp. Phys., 190 (2003), 478-500. doi: 10.1016/S0021-9991(03)00294-8
|
| [16] |
K. J. Painter, P. K. Maini, and H. G. Othmer, Stripe formation in juvenile Pomacanthus explained by a generalized Turing mechanism with chemotaxis, Proc. Nat. Acad. Sci., 96 (1999), 5549-5554. doi: 10.1073/pnas.96.10.5549
|
| [17] |
S. S. Lee, E. A. Gaffney, and R. E. Baker, The dynamics of Turing patterns for morphogenregulated growing domains with cellular response delays, Bull. Math. Biol., 73 (2011), 2527-2551. doi: 10.1007/s11538-011-9634-8
|
| [18] | J. D. Murray, Mathematical Biology: I. An Introduction, volume 17. Springer Science & Business Media, 2007. |
| [19] | B. Ermentrout, Stripes or spots? Nonlinear effects in bifurcation of reaction-diffusion equations on the square, Proc. R. Soc. A, 434 (1991), 413-417. |
| [20] |
H. Shoji, Y. Iwasa, and S. Kondo, Stripes, spots, or reversed spots in two-dimensional Turing systems, J. Theor. Biol., 224 (2003), 339-350. doi: 10.1016/S0022-5193(03)00170-X
|
| [21] | H. Meinhardt, Tailoring and coupling of reaction-diffusion systems to obtain reproducible complex pattern formation during development of the higher organisms, Appl. Math. Comput., 32 (1989), 103-135. |
| [22] |
R. Chaturvedi, C. Huang, B. Kazmierczak, T. Schneider, J. A. Izaguirre, T. Glimm, H. G. E. Hentschel, J. A. Glazier, S. A. Newman and M. S. Alber, On multiscale approaches to three-dimensional modelling of morphogenesis, J. R. Soc. Interface, 2 (2005), 237-253. doi: 10.1098/rsif.2005.0033
|
| [23] |
S. Christley, M. S. Alber and S. A. Newman, Patterns of mesenchymal condensation in a multiscale, discrete stochastic model, PLoS Comput. Biol., 3 (2007), e76. doi: 10.1371/journal.pcbi.0030076
|
| [24] | B. Duggan and J. Metzcar, Generating Turing-like patterns in an off-lattice agent based model: Handout, Preprint. |
| [25] |
D. Karig, K. M. Martini, T. Lu, N. A. DeLateur, N. Goldenfeld, and R. Weiss, Stochastic Turing patterns in a synthetic bacterial population, Proc. Nat. Acad. Sci., 115 (2018), 6572-6577. doi: 10.1073/pnas.1720770115
|
| [26] |
M. A. Kiskowski, M. S. Alber, G. L. Thomas, J. A. Glazier, N. B. Bronstein, J. Pu and S. A. Newman, Interplay between activator-inhibitor coupling and cell-matrix adhesion in a cellular automaton model for chondrogenic patterning, Dev. Biol., 271 (2004), 372-387. doi: 10.1016/j.ydbio.2004.03.038
|
| [27] | S. Kondo, Turing pattern formation without diffusion, in Conference on Computability in Europe, Springer, 2012,416-421. |
| [28] |
J. Moreira and A. Deutsch, Pigment pattern formation in zebrafish during late larval stages: A model based on local interactions, Dev. Dyn., 232 (2005), 33-42. doi: 10.1002/dvdy.20199
|
| [29] |
S. Okuda, T. Miura, Y. Inoue, T. Adachi, and M. Eiraku, Combining Turing and 3D vertex models reproduces autonomous multicellular morphogenesis with undulation, tubulation, and branching, Sci. Rep., 8 (2018), 1-15. doi: 10.1038/s41598-017-17765-5
|
| [30] |
A. Volkening and B. Sandstede, Modelling stripe formation in zebrafish: An agent-based approach, J. R. Soc. Interface, 12 (2015), 20150812. doi: 10.1098/rsif.2015.0812
|
| [31] |
C. M. Glen, M. L. Kemp, and E. O. Voit, Agent-based modeling of morphogenetic systems: Advantages and challenges, PLoS Comp. Biol., 15 (2019), e1006577. doi: 10.1371/journal.pcbi.1006577
|
| [32] |
J. A. Izaguirre, R. Chaturvedi, C. Huang, T. Cickovski, J. Coffland, G. Thomas, G. Forgacs, M. Alber, G. Hentschel, S. A. Newman, et al, CompuCell, a multi-model framework for simulation of morphogenesis, Bioinformatics, 20 (2004), 1129-1137. doi: 10.1093/bioinformatics/bth050
|
| [33] |
T. M. Cickovski, C. Huang, R. Chaturvedi, T. Glimm, H. G. E. Hentschel, M. S. Alber, J. A. Glazier, S. A. Newman, and J. A. Izaguirre, A framework for three-dimensional simulation of morphogenesis, Trans. Comput. Biol. Bioinform., 2 (2005), 273-288. doi: 10.1109/TCBB.2005.46
|
| [34] | B. D. Hughes, Random walks and random environments: Random walks, vol. 1, Oxford University Press, 1995. |
| [35] |
T. Hillen and H. G. Othmer, The diffusion limit of transport equations derived from velocity-jump processes, SIAM J. Appl. Math., 61 (2000), 751-775. doi: 10.1137/S0036139999358167
|
| [36] |
T. Hillen and K. J. Painter, A user's guide to PDE models for chemotaxis, J. Math. Biol., 58 (2009), 183. doi: 10.1007/s00285-008-0201-3
|
| [37] |
H. G. Othmer, S. R. Dunbar and W. Alt, Models of dispersal in biological systems, J. Math. Biol., 26 (1988), 263-298. doi: 10.1007/BF00277392
|
| [38] | K. J. Painter and T. Hillen, Volume-filling and quorum-sensing in models for chemosensitive movement, Can. Appl. Math. Quart., 10. (2002), 501-543. |
| [39] |
K. J. Painter and J. A. Sherratt, Modelling the movement of interacting cell populations, J. Theor. Biol., 225 (2003), 327-339. doi: 10.1016/S0022-5193(03)00258-3
|
| [40] |
W. Alt, Biased random walk models for chemotaxis and related diffusion approximations, J. Math. Biol., 9 (1980), 147-177. doi: 10.1007/BF00275919
|
| [41] |
M. Burger, P. Markowich and J.-F. Pietschmann, Continuous limit of a crowd motion and herding model: analysis and numerical simulations, Kinet. Relat. Models, 4 (2011), 1025-1047. doi: 10.3934/krm.2011.4.1025
|
| [42] |
A. Stevens, The derivation of chemotaxis equations as limit dynamics of moderately interacting stochastic many-particle systems, SIAM J. Appl. Math, 61 (2000), 183-212. doi: 10.1137/S0036139998342065
|
| [43] |
A. Stevens and H. G. Othmer, Aggregation, blowup, and collapse: the abc's of taxis in reinforced random walks, SIAM J. Appl. Math., 57 (1997), 1044-1081. doi: 10.1137/S0036139995288976
|
| [44] |
F. Bubba, T. Lorenzi, and F. R. Macfarlane, From a discrete model of chemotaxis with volume-filling to a generalized patlak-keller-segel model, Proc. R. Soc. A, 476 (2020), 20190871. doi: 10.1098/rspa.2019.0871
|
| [45] |
N. Champagnat and S. Méléard, Invasion and adaptive evolution for individual-based spatially structured populations, J. Math. Biol., 55 (2007), 147. doi: 10.1007/s00285-007-0072-z
|
| [46] | M. A. J. Chaplain, T. Lorenzi and F. R. Macfarlane, Bridging the gap between individual-based and continuum models of growing cell populations, J. Math. Biol., 80 (2020), 342-371. |
| [47] |
M. Inoue, Derivation of a porous medium equation from many markovian particles and the propagation of chaos, Hiroshima Math. J., 21 (1991), 85-110. doi: 10.32917/hmj/1206128924
|
| [48] |
K. Oelschläger, On the derivation of reaction-diffusion equations as limit dynamics of systems of moderately interacting stochastic processes, Probab. Theory Relat. Fields, 82 (1989), 565-586. doi: 10.1007/BF00341284
|
| [49] |
H. G. Othmer and T. Hillen, The diffusion limit of transport equations Ⅱ: Chemotaxis equations, SIAM J. Appl. Math., 62 (2002), 1222-1250. doi: 10.1137/S0036139900382772
|
| [50] |
C. J. Penington, B. D. Hughes and K. A. Landman, Building macroscale models from microscale probabilistic models: A general probabilistic approach for nonlinear diffusion and multispecies phenomena, Phys. Rev. E, 84 (2011), 041120. doi: 10.1103/PhysRevE.84.041120
|
| [51] | C. J. Penington, B. D. Hughes and K. A. Landman, Interacting motile agents: Taking a mean-field approach beyond monomers and nearest-neighbor steps, Phys. Rev. E, 89 (2014), 032714. |
| [52] |
R. E. Baker, A. Parker and M. J. Simpson, A free boundary model of epithelial dynamics, J. Theor. Biol., 481 (2019), 61-74. doi: 10.1016/j.jtbi.2018.12.025
|
| [53] |
H. M. Byrne and D. Drasdo, Individual-based and continuum models of growing cell populations: A comparison, J. Math. Biol., 58 (2009), 657. doi: 10.1007/s00285-008-0212-0
|
| [54] |
T. Lorenzi, P. J. Murray and M. Ptashnyk, From individual-based mechanical models of multicellular systems to free-boundary problems, Interface Free Bound., 22 (2020), 205-244. doi: 10.4171/IFB/439
|
| [55] |
S. Motsch and D. Peurichard, From short-range repulsion to hele-shaw problem in a model of tumor growth, J. Math. Biol., 76 (2018), 205-234. doi: 10.1007/s00285-017-1143-4
|
| [56] |
P. J. Murray, C. M. Edwards, M. J. Tindall and P. K. Maini, From a discrete to a continuum model of cell dynamics in one dimension, Phys. Rev. E, 80 (2009), 031912. doi: 10.1103/PhysRevE.80.031912
|
| [57] |
P. J. Murray, C. M. Edwards, M. J. Tindall and P. K. Maini, Classifying general nonlinear force laws in cell-based models via the continuum limit, Phys. Rev. E, 85 (2012), 021921. doi: 10.1103/PhysRevE.85.021921
|
| [58] |
K. Oelschläger, Large systems of interacting particles and the porous medium equation, J. Diff. equation., 88 (1990), 294-346. doi: 10.1016/0022-0396(90)90101-T
|
| [59] |
B. J. Binder and K. A. Landman, Exclusion processes on a growing domain, J. Theor. Biol., 259 (2009), 541-551. doi: 10.1016/j.jtbi.2009.04.025
|
| [60] |
L. Dyson, P. K. Maini and R. E. Baker, Macroscopic limits of individual-based models for motile cell populations with volume exclusion, Phys. Rev. E, 86 (2012), 031903. doi: 10.1103/PhysRevE.86.031903
|
| [61] |
A. E. Fernando, K. A. Landman and M. J. Simpson, Nonlinear diffusion and exclusion processes with contact interactions, Phys. Rev. E, 81 (2010), 011903. doi: 10.1103/PhysRevE.81.011903
|
| [62] |
S. T. Johnston, R. E. Baker, D. S. McElwain and M. J. Simpson, Co-operation, competition and crowding: a discrete framework linking allee kinetics, nonlinear diffusion, shocks and sharp-fronted travelling waves, Sci. Rep., 7 (2017), 42134. doi: 10.1038/srep42134
|
| [63] |
S. T. Johnston, M. J. Simpson and R. E. Baker, Mean-field descriptions of collective migration with strong adhesion, Phys. Rev. E, 85 (2012), 051922. doi: 10.1103/PhysRevE.85.051922
|
| [64] |
K. A. Landman and A. E. Fernando, Myopic random walkers and exclusion processes: Single and multispecies, Phys. A Stat. Mech. Appl., 390 (2011), 3742-3753. doi: 10.1016/j.physa.2011.06.034
|
| [65] |
P. M. Lushnikov, N. Chen and M. Alber, Macroscopic dynamics of biological cells interacting via chemotaxis and direct contact, Phys. Rev. E, 78 (2008), 061904. doi: 10.1103/PhysRevE.78.061904
|
| [66] |
M. J. Simpson, K. A. Landman and B. D. Hughes, Cell invasion with proliferation mechanisms motivated by time-lapse data, Phys. A Stat. Mech. Appl., 389 (2010), 3779-3790. doi: 10.1016/j.physa.2010.05.020
|
| [67] |
C. Deroulers, M. Aubert, M. Badoual and B. Grammaticos, Modeling tumor cell migration: from microscopic to macroscopic models, Phys. Rev. E, 79 (2009), 031917. doi: 10.1103/PhysRevE.79.031917
|
| [68] |
D. Drasdo, Coarse graining in simulated cell populations, Adv. Complex Syst., 8 (2005), 319-363. doi: 10.1142/S0219525905000440
|
| [69] |
M. J. Simpson, A. Merrifield, K. A. Landman and B. D. Hughes, Simulating invasion with cellular automata: connecting cell-scale and population-scale properties, Phys. Rev. E, 76 (2007), 021918. doi: 10.1103/PhysRevE.76.021918
|
| [70] |
A. Buttenschoen, T. Hillen, A. Gerisch and K. J. Painter, A space-jump derivation for non-local models of cell-cell adhesion and non-local chemotaxis, J. Math. Biol., 76 (2018), 429-456. doi: 10.1007/s00285-017-1144-3
|
| [71] |
L. Marcon and J. Sharpe, Turing patterns in development: What about the horse part? Curr. Opin. Genet. Dev., 22 (2012), 578-584. doi: 10.1016/j.gde.2012.11.013
|
| [72] | H. G. Othmer, P. K. Maini and J. D. Murray, Experimental and theoretical advances in biological pattern formation, vol. 259, Springer Science & Business Media, 2012. |
| [73] |
H. G. Othmer, K. J. Painter, D. Umulis, and C. Xue, The intersection of theory and application in elucidating pattern formation in developmental biology, Math. Model. Nat. Phenom., 4 (2009), 3-82. doi: 10.1051/mmnp/20094401
|
| [74] |
P. Maini, K. Painter and H. P. Chau, Spatial pattern formation in chemical and biological systems, J. Chem. Soc. Faraday Trans., 93 (1997), 3601-3610. doi: 10.1039/a702602a
|
| [75] |
M. R. Myerscough, P. K. Maini, and K. J. Painter, Pattern formation in a generalized chemotactic model, Bull. Math. Biol., 60 (1998), 1-26. doi: 10.1006/bulm.1997.0010
|
| [76] |
J. A. Sherratt and J. D. Murray, Mathematical analysis of a basic model for epidermal wound healing, J. Math. Biol., 29 (1991), 389-404. doi: 10.1007/BF00160468
|
| [77] |
J. Schnakenberg, Simple chemical reaction systems with limit cycle behaviour, J. Theor. Biol., 81 (1979), 389-400. doi: 10.1016/0022-5193(79)90042-0
|
| [78] | G. Lolas, Spatio-temporal pattern formation and reaction diffusion systems, Master's thesis, University of Dundee Scotland, 1999. |
| [79] |
R. McLennan, L. Dyson, K. W. Prather, J. A. Morrison, R. E. Baker, P. K. Maini, and P. M. Kulesa, Multiscale mechanisms of cell migration during development: Theory and experiment, Development, 139 (2012), 2935-2944. doi: 10.1242/dev.081471
|
| [80] |
R. McLennan, L. J. Schumacher, J. A. Morrison, J. M. Teddy, D. A. Ridenour, A. C. Box, C. L. Semerad, H. Li, W. McDowell, D. Kay, et al, Neural crest migration is driven by a few trailblazer cells with a unique molecular signature narrowly confined to the invasive front, Development, 142 (2015), 2014-2025. doi: 10.1242/dev.117507
|
| [81] |
R. McLennan, L. J. Schumacher, J. A. Morrison, J. M. Teddy, D. A. Ridenour, A. C. Box, C. L. Semerad, H. Li, W. McDowell, D. Kay, et al, VEGF signals induce trailblazer cell identity that drives neural crest migration, Dev. Biol., 407 (2015), 12-25. doi: 10.1016/j.ydbio.2015.08.011
|
| [82] | J. D. Murray, G. F. Oster, and A. K. Harris, A mechanical model for mesenchymal morphogenesis, J. Math. Biol., 17 (1983), 125-129. |
| [83] |
F. Schweisguth and F. Corson, Self-organization in pattern formation, Develop. Cell, 49 (2019), 659-677. doi: 10.1016/j.devcel.2019.05.019
|
| [84] |
L. Tweedy, D. A. Knecht, G. M. Mackay, and R. H. Insall, Self-generated chemoattractant gradients: attractant depletion extends the range and robustness of chemotaxis, PLoS Biol., 14 (2016), e1002404. doi: 10.1371/journal.pbio.1002404
|
| [85] |
J. Galle, M. Loeffler, and D. Drasdo, Modeling the effect of deregulated proliferation and apoptosis on the growth dynamics of epithelial cell populations in vitro, Biophys. J., 88 (2005), 62-75. doi: 10.1529/biophysj.104.041459
|
| [86] | J. Galle, G. Aust, G. Schaller, T. Beyer, and D. Drasdo, Individual cell-based models of the spatial-temporal organization of multicellular systems-Achievements and limitations, Cytom. A, 69 (2006), 704-710. |
| [87] |
D. Drasdo, S. Hoehme, and M. Block, On the role of physics in the growth and pattern formation of multi-cellular systems: What can we learn from individual-cell based models? J. Stat. Phys., 128 (2007), 287. doi: 10.1007/s10955-007-9289-x
|
| [88] |
M. P. Neilson, D. M. Veltman, P. J. M. van Haastert, S. D. Webb, J. A. Mackenzie, and R. H. Insall, Chemotaxis: A feedback-based computational model robustly predicts multiple aspects of real cell behaviour, PLoS Biol., 9 (2011), e1000618. doi: 10.1371/journal.pbio.1000618
|
| [89] |
S. Kondo, An updated kernel-based turing model for studying the mechanisms of biological pattern formation, J. Theor Biol., 414 (2017), 120-127. doi: 10.1016/j.jtbi.2016.11.003
|
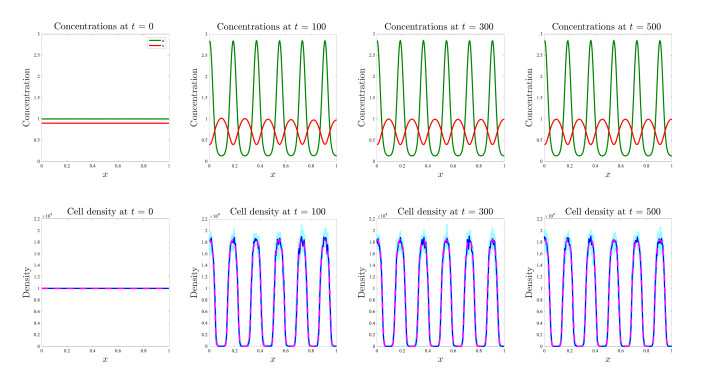
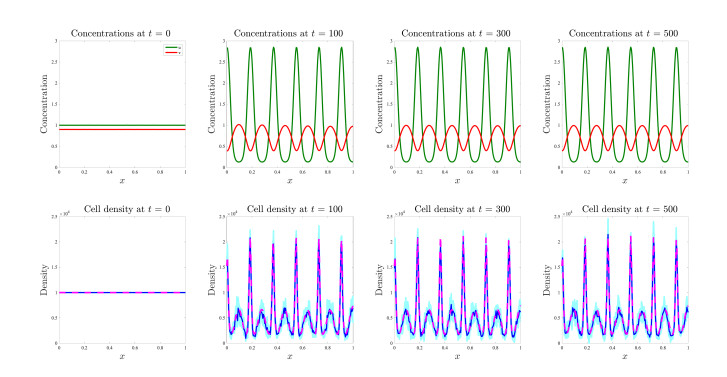
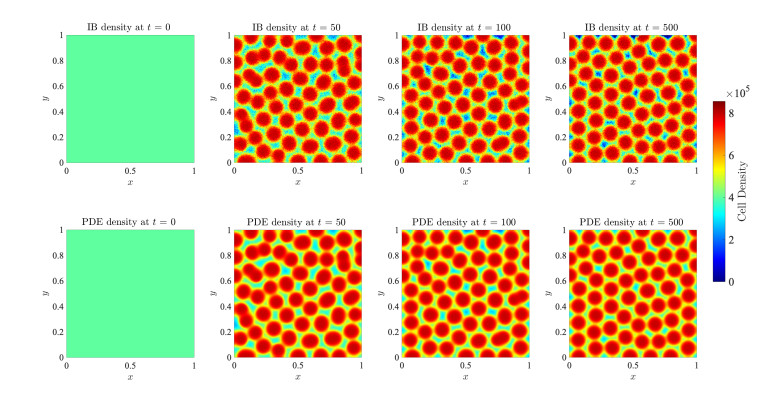
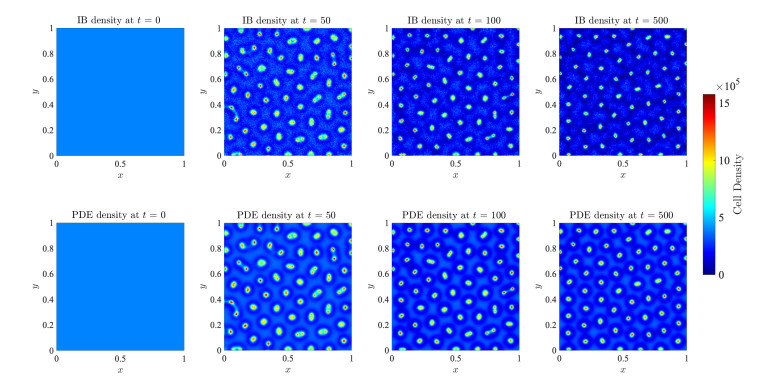
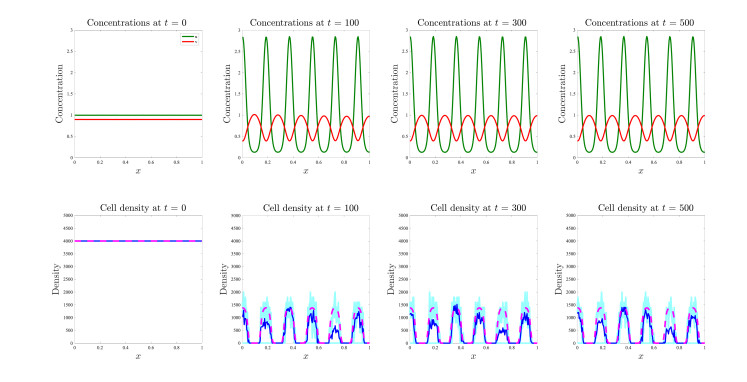
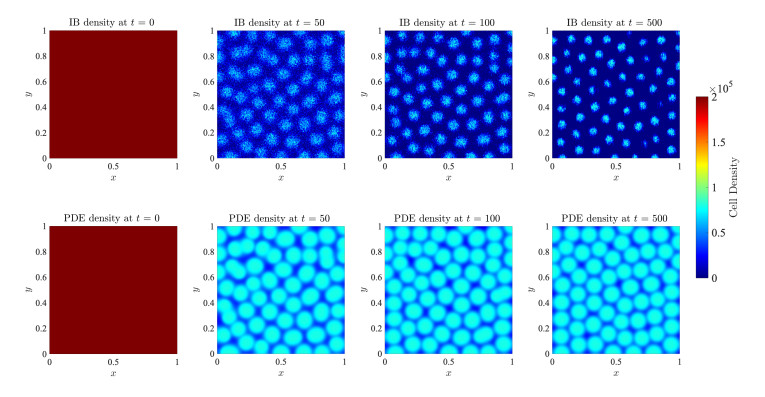
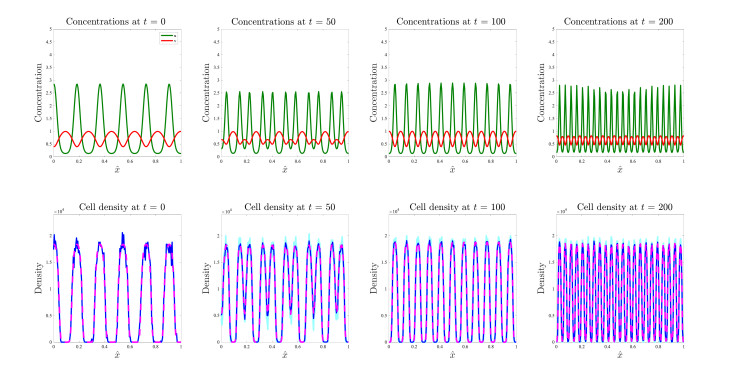
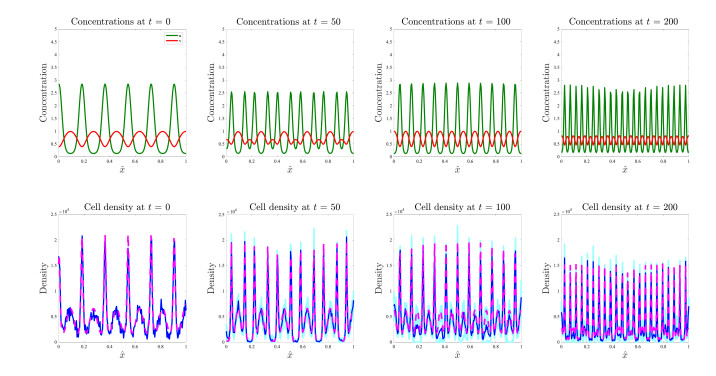

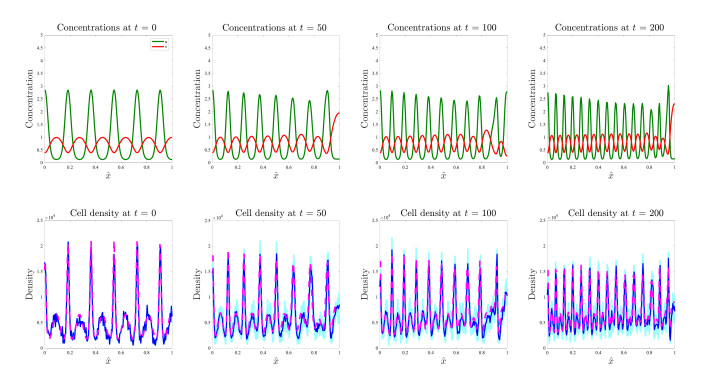
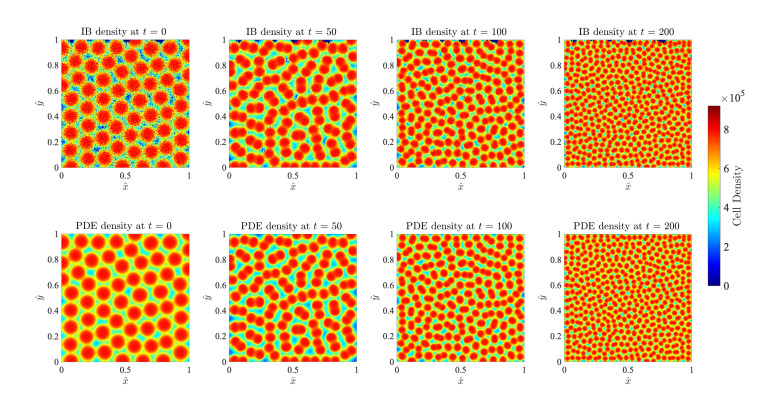
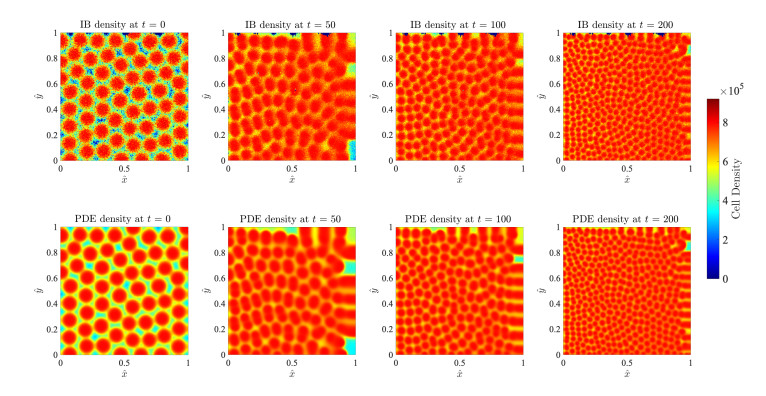
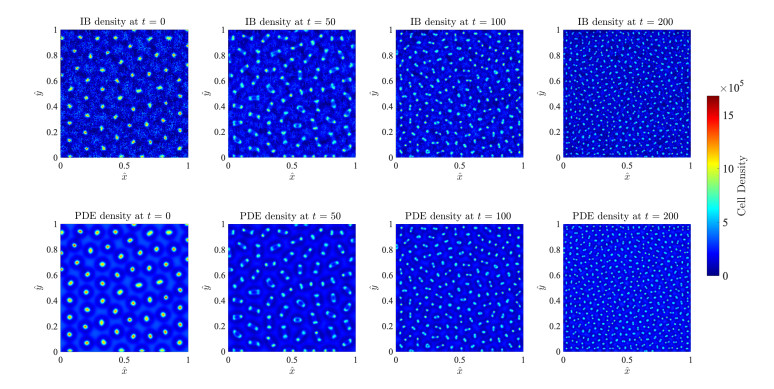
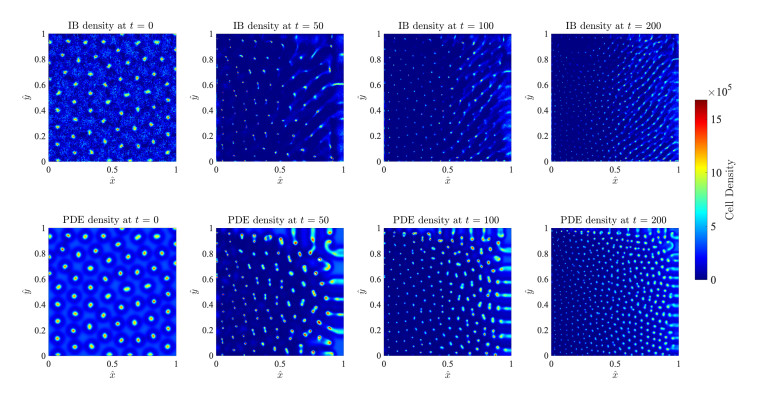
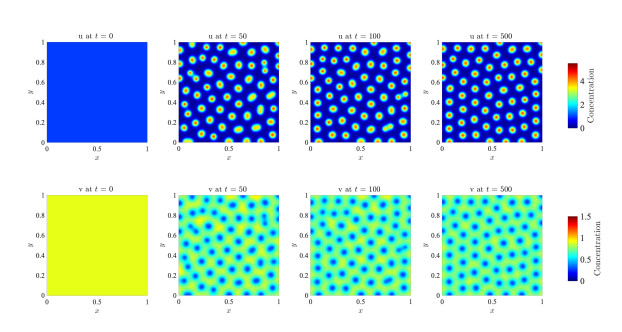
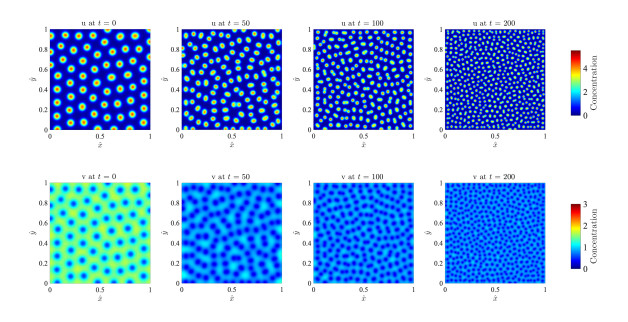
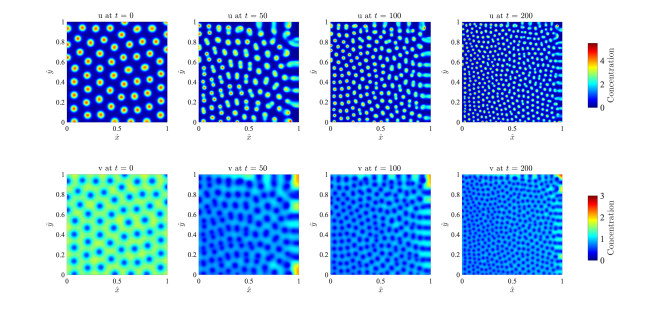
Figures(17)

Fiona R. Macfarlane, Mark A. J. Chaplain, Tommaso Lorenzi. A hybrid discrete-continuum approach to model Turing pattern formation[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7442-7479. doi: 10.3934/mbe.2020381







 DownLoad:
DownLoad: