Citation: Agustín Gabriel Yabo, Jean-Baptiste Caillau, Jean-Luc Gouzé. Optimal bacterial resource allocation: metabolite production in continuous bioreactors[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7074-7100. doi: 10.3934/mbe.2020364

| [1] | R. U. Ibarra, J. S. Edwards, B. O. Palsson, Escherichia coli k-12 undergoes adaptive evolution to achieve in silico predicted optimal growth, Nature, 420 (2002), 186. |
| [2] |
E. Bosdriesz, D. Molenaar, B. Teusink, F. J. Bruggeman, How fast-growing bacteria robustly tune their ribosome concentration to approximate growth-rate maximization, FEBS J., 282 (2015), 2029-2044. doi: 10.1111/febs.13258

|
| [3] |
J. D. Van Elsas, A. V. Semenov, R. Costa, J. T. Trevors, Survival of escherichia coli in the environment: fundamental and public health aspects, ISME J., 5 (2011), 173-183. doi: 10.1038/ismej.2010.80
|
| [4] | N. Giordano, F. Mairet, J.-L. Gouzé, J. Geiselmann, H. De Jong, Dynamical allocation of cellular resources as an optimal control problem: novel insights into microbial growth strategies, PLoS Comput. Biol., 12 (2016), e1004802. |
| [5] | H. De Jong, S. Casagranda, N. Giordano, E. Cinquemani, D. Ropers, J. Geiselmann, et al., Mathematical modelling of microbes: metabolism, gene expression and growth, J. R. Soc. Interface, 14 (2017), 20170502. |
| [6] | A. L. Koch, Why can't a cell grow infinitely fast?, Can. J. Microbiol., 34 (1988), 421-426. |
| [7] |
J. Shu, M. Shuler, A mathematical model for the growth of a single cell of E. coli on a glucose/glutamine/ammonium medium, Biotechnol. Bioeng., 33 (1989), 1117-1126. doi: 10.1002/bit.260330907
|
| [8] | D. V. Goeddel, D. G. Kleid, F. Bolivar, H. L. Heyneker, D. G. Yansura, R. Crea, et al., Expression in escherichia coli of chemically synthesized genes for human insulin, Proc. Natl. Acad. Sci., 76 (1979), 106-110. |
| [9] |
L. Huo, J. J. Hug, C. Fu, X. Bian, Y. Zhang, R. Müller, Heterologous expression of bacterial natural product biosynthetic pathways, Nat. Product Rep., 36 (2019), 1412-1436. doi: 10.1039/C8NP00091C
|
| [10] | I. Yegorov, F. Mairet, H. De Jong, J.-L. Gouzé, Optimal control of bacterial growth for the maximization of metabolite production, J. Math. Biol., 78 (2019), 985-1032. |
| [11] | E. Cinquemani, F. Mairet, I. Yegorov, H. de Jong, J.-L. Gouzé, Optimal control of bacterial growth for metabolite production: the role of timing and costs of control, in 2019 18th European Control Conference (ECC), IEEE, 2019, 2657-2662. |
| [12] | A. G. Yabo, J.-B. Caillau, J.-L. Gouzé, Singular regimes for the maximization of metabolite production, in 2019 IEEE 58th Conference on Decision and Control (CDC), IEEE, 2019, 31-36. |
| [13] | J. Izard, C. D. G. Balderas, D. Ropers, S. Lacour, X. Song, Y. Yang, et al., A synthetic growth switch based on controlled expression of rna polymerase, Mol. Syst. Biol., 11 (2015), 840. |
| [14] | I. Otero-Muras, A. A. Mannan, J. R. Banga, D. A. Oyarzún, Multiobjective optimization of gene circuits for metabolic engineering, IFAC-PapersOnLine, 52 (2019), 13-16. |
| [15] | D. Molenaar, R. Van Berlo, D. De Ridder, B. Teusink, Shifts in growth strategies reflect tradeoffs in cellular economics, Mol. Syst. Biol., 5 (2009), 323. |
| [16] | M. T. Wortel, E. Bosdriesz, B. Teusink, F. J. Bruggeman, Evolutionary pressures on microbial metabolic strategies in the chemostat, Sci. Rep., 6 (2016), 29503. |
| [17] |
D. W. Spitzer, Maximization of steady-state bacterial production in a chemostat with ph and substrate control, Biotechnol. Bioeng., 18 (1976), 167-178. doi: 10.1002/bit.260180203
|
| [18] |
R. R. Lichtl, M. J. Bazin, D. O. Hall, The biotechnology of hydrogen production by nostoc flagelliforme grown under chemostat conditions, Appl. Microbiol. Biotechnol., 47 (1997), 701-707. doi: 10.1007/s002530050998
|
| [19] |
M. C. D'anjou, A. J. Daugulis, A rational approach to improving productivity in recombinant pichia pastoris fermentation, Biotechnol. Bioeng., 72 (2001), 1-11. doi: 10.1002/1097-0290(20010105)72:1<1::AID-BIT1>3.0.CO;2-T
|
| [20] |
T. Bayen, F. Mairet, Optimization of the separation of two species in a chemostat, Automatica, 50 (2014), 1243-1248. doi: 10.1016/j.automatica.2014.02.024
|
| [21] |
C. Martínez, O. Bernard, F. Mairet, Maximizing microalgae productivity in a light-limited chemostat, IFAC-PapersOnLine, 51 (2018), 735-740. doi: 10.1016/j.ifacol.2018.05.070
|
| [22] | H. R. Thieme, Convergence results and a PoincaréBendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755-763. |
| [23] | L. S. Pontryagin, Mathematical Theory of Optimal Processes, Routledge, 2018. |
| [24] | H. L. Smith, P. Waltman, The Theory of the Chemostat: Dynamics of Microbial Competition, Cambridge University Press, 1995. |
| [25] | H. Bremer, P. P. Dennis, Modulation of chemical composition and other parameters of the cell by growth rate, in Escherichia coli and Salmonella: Cellular and Molecular Biology, ASM Press, Washington, (1996), 1553-1569. |
| [26] | S. Goutelle, M. Maurin, F. Rougier, X. Barbaut, L. Bourguignon, M. Ducher, et al., The hill equation: a review of its capabilities in pharmacological modelling, Fundam. Clin. Pharmacol., 22 (2008), 633-648. |
| [27] |
K. A. Johnson, R. S. Goody, The original Michaelis constant: translation of the 1913 Michaelis-Menten paper, Biochemistry, 50 (2011), 8264-8269. doi: 10.1021/bi201284u
|
| [28] | A. A. Agrachev, Y. Sachkov, Control Theory from the Geometric Viewpoint, Springer Science & Business Media, 2013. |
| [29] | M. I. Zelikin, V. F. Borisov, Theory of Chattering Control: With Applications to Astronautics, Robotics, Economics, and Engineering, Springer Science & Business Media, 2012. |
| [30] | A. Van der Schaft, Symmetries in optimal control, SIAM J. Control Optim., 25 (1987), 245-259. |
| [31] |
E. Trélat, E. Zuazua, The turnpike property in finite-dimensional nonlinear optimal control, J. Differ. Equations, 258 (2015), 81-114. doi: 10.1016/j.jde.2014.09.005
|
| [32] | W. Djema, L. Giraldi, S. Maslovskaya, O. Bernard, Turnpike features in optimal selection of species represented by quota models, Submitted. |
| [33] | I. S. Team Commands, Bocop: an open source toolbox for optimal control, 2017. Available from: http://bocop.org. |
| [34] |
H. Robbins, A generalized Legendre-Clebsch condition for the singular cases of optimal control, IBM J. Res. Dev., 11 (1967), 361-372. doi: 10.1147/rd.114.0361
|
| [35] |
M. Scott, C. W. Gunderson, E. M. Mateescu, Z. Zhang, T. Hwa, Interdependence of cell growth and gene expression: origins and consequences, Science, 330 (2010), 1099-1102. doi: 10.1126/science.1192588
|
| [36] | I. Yegorov, F. Mairet, J.-L. Gouzé, Optimal feedback strategies for bacterial growth with degradation, recycling, and effect of temperature, Optim. Control Appl. Methods, 39 (2018), 1084-1109. |
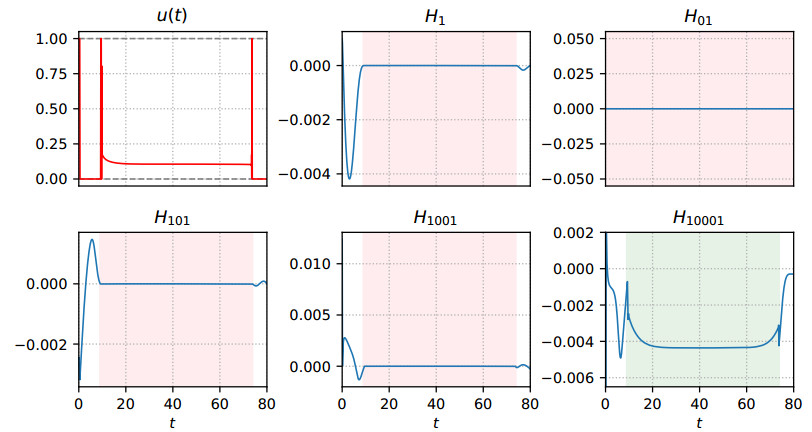

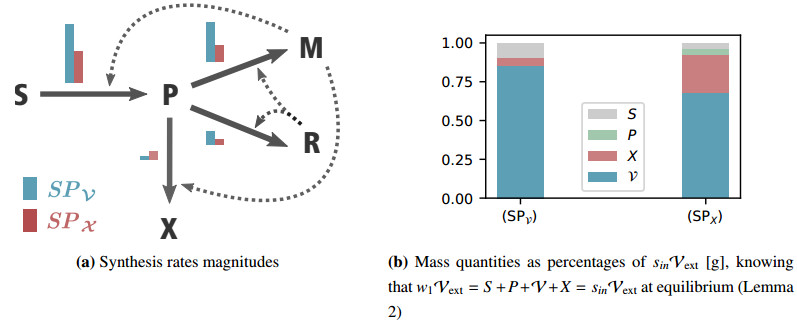
Figures(7) / Tables(1)

Agustín Gabriel Yabo, Jean-Baptiste Caillau, Jean-Luc Gouzé. Optimal bacterial resource allocation: metabolite production in continuous bioreactors[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7074-7100. doi: 10.3934/mbe.2020364







 DownLoad:
DownLoad: