Citation: Yuyi Xue, Yanni Xiao. Analysis of a multiscale HIV-1 model coupling within-host viral dynamics and between-host transmission dynamics[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6720-6736. doi: 10.3934/mbe.2020350

| [1] | R. M. Anderson, R. M. May, O. U. P. (OUP), Infectious diseases of human: dynamics and control, 1992. |
| [2] | O. Diekmann, J. Heesterbeek, Mathematical epidemiology of infectious diseases: Model building, analysis and interpretation, Wiley Series in Mathematical and Computational Biology, Chichester, Wiley. |
| [3] |
A. S. Perelson, P. W. Nelson, Mathematical Analysis Of HIV-1 Dynamics In Vivo, Siam. Rev., 41 (1999), 3-44. doi: 10.1137/S0036144598335107

|
| [4] |
P. Song, Y. Lou, Y. Xiao, A spatial seirs reaction-diffusion model in heterogeneous environment, J. Differ. Equations, 267 (2019), 5084-5114. doi: 10.1016/j.jde.2019.05.022
|
| [5] | S. Wain-Hobson, Virus dynamics: Mathematical principles of immunology and virology, Nat. Med., 410 (2001), 412-413. |
| [6] | L. J. Abu-Raddad, R. V. Barnabas, H. Janes, H. A. Weiss, J. G. Kublin, I. M. Longini, et al., Have the explosive HIV epidemics in sub-Saharan Africa been driven by higher community viral load?, AIDS, 27 (2013), 2494-2496. |
| [7] |
D. Wilson, M. Law, A. E. Grulich, D. A. Cooper, J. M. Kaldor, Relation between HIV viral load and infectiousness: a model-based analysis, The Lancet, 372 (2008), 314-320. doi: 10.1016/S0140-6736(08)61115-0
|
| [8] |
T. C. Quinn, M. J. Wawer, N. Sewankambo, D. Serwadda, R. H. Gray, Viral load and heterosexual transmission of human immunodeficiency virus type 1, New. Engl. J. Med., 342 (2000), 921-929. doi: 10.1056/NEJM200003303421303
|
| [9] | L. M. Childs, F. E. Moustaid, Z. Gajewski, S. Kadelka, R. Nikinbeers, J. W. Smith, et al., Linked within-host and between-host models and data for infectious diseases: a systematic review, PeerJ, 7 (2019), e7057. |
| [10] | N. Dorratoltaj, R. Nikinbeers, S. M. Ciupe, S. Eubank, K. Abbas, Multi-scale immunoepidemiological modeling of within-host and between-host HIV dynamics: systematic review of mathematical models, PeerJ, 5 (2017), e3877. |
| [11] |
A. Gandolfi, A. Pugliese, C. Sinisgalli, Epidemic dynamics and host immune response: a nested approach, J. Math. Biol., 70 (2015), 399-435. doi: 10.1007/s00285-014-0769-8
|
| [12] | W. Garira, A primer on multiscale modelling of infectious disease systems, Infect. Dis. Model., 3 (2018), 176-191. |
| [13] |
W. Garira, A complete categorization of multiscale models of infectious disease systems, J. Biol. Dyn., 11 (2017), 378-435. doi: 10.1080/17513758.2017.1367849
|
| [14] | M. A. Gilchrist, D. Coombs, Evolution of virulence: Interdependence, constraints, and selection using nested models, Theor. Popul. Biol., 69 (2006), 145-153. |
| [15] |
M. Park, C. Loverdo, S. J. Schreiber, J. O. Lloydsmith, Multiple scales of selection influence the evolutionary emergence of novel pathogens, Philos. T. R. Soc. B., 368 (2013), 20120333. doi: 10.1098/rstb.2012.0333
|
| [16] |
M. Shen, Y. Xiao, L. Rong, Global stability of an infection-age structured HIV-1 model linking within-host and between-host dynamics, Math. Biosci., 263 (2015), 37-50. doi: 10.1016/j.mbs.2015.02.003
|
| [17] |
M. Shen, Y. Xiao, L. Rong, L. A. Meyers, Conflict and accord of optimal treatment strategies for HIV infection within and between hosts, Math. Biosci., 309 (2019), 107-117. doi: 10.1016/j.mbs.2019.01.007
|
| [18] |
M. Shen, Y. Xiao, L. Rong, G. Zhuang, Global dynamics and cost-effectiveness analysis of HIV pre-exposure prophylaxis and structured treatment interruptions based on a multi-scale model, Appl. Math. Model., 75 (2019), 162-200. doi: 10.1016/j.apm.2019.05.024
|
| [19] | Z. Feng, X. Cen, Y. Zhao, J. Velasco-Hernandez, Coupled within-host and between-host dynamics and evolution of virulence, Math. Biosci., 270 (2015), 204-212. |
| [20] | Z. Feng, J. Velasco-Hernandez, B. Tapia-Santos, A mathematical model for coupling within-host and between-host dynamics in an environmentally-driven infectious disease, Math. Biosci., 241 (2013), 49-55. |
| [21] |
X. Wang, S. Tang, A multiscale model on hospital infections coupling macro and micro dynamics, Commun. Nonlinear. Sci., 50 (2017), 256-270. doi: 10.1016/j.cnsns.2017.03.009
|
| [22] |
X. Sun, Y. Xiao, Multiscale system for environmentally-driven infectious disease with threshold control strategy, Int. J. Bifurcat. Chaos, 28 (2018), 1850064. doi: 10.1142/S0218127418500645
|
| [23] |
Y. Xiao, C. Xiang, R. Cheke, S. Tang, Coupling the macroscale to the microscale in a spatiotemporal context to examine effects of spatial diffusion on disease transmission, B. Math. Biol., 82 (2020), 1-27. doi: 10.1007/s11538-019-00680-3
|
| [24] |
S. Bhattacharya, M. Martcheva, An immuno-eco-epidemiological model of competition, J. Biol. Dyn., 10 (2016), 314-341. doi: 10.1080/17513758.2016.1186291
|
| [25] |
T. Kostova, Persistence of viral infections on the population level explained by an immunoepidemiological model, Math. Biosci., 206 (2007), 309-319. doi: 10.1016/j.mbs.2005.08.003
|
| [26] |
E. C. Manda, F. Chirove, Modelling coupled within host and population dynamics of and HIV infection, J. Math. Biol., 76 (2018), 1123-1158. doi: 10.1007/s00285-017-1170-1
|
| [27] |
X. Cen, Z. Feng, Y. Zhao, Emerging disease dynamics in a model coupling within-host and between-host systems, J. Theor. Biol., 361 (2014), 141-151. doi: 10.1016/j.jtbi.2014.07.030
|
| [28] |
B. Boldin, O. Diekmann, Superinfections can induce evolutionarily stable coexistence of pathogens, J. Math. Biol., 56 (2008), 635-672. doi: 10.1007/s00285-007-0135-1
|
| [29] |
P. Dreessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
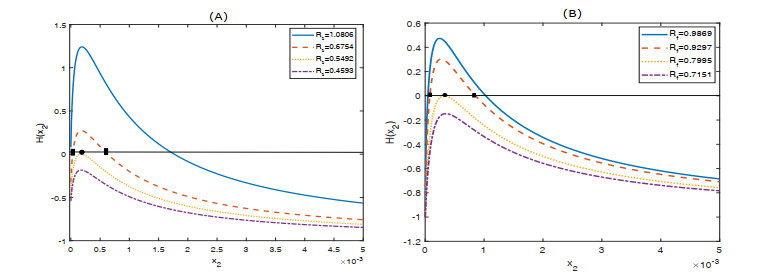
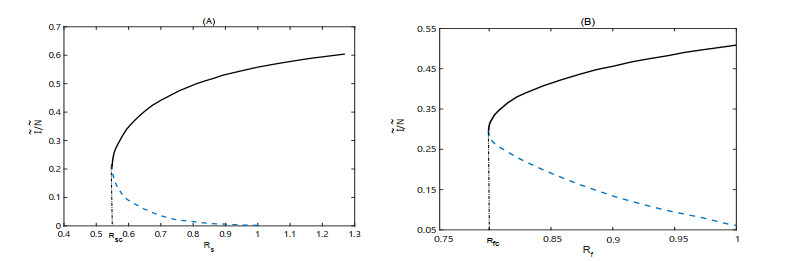
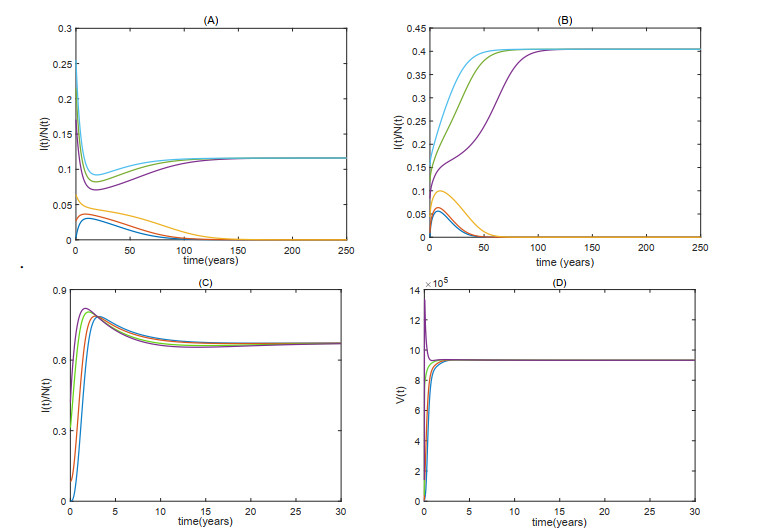
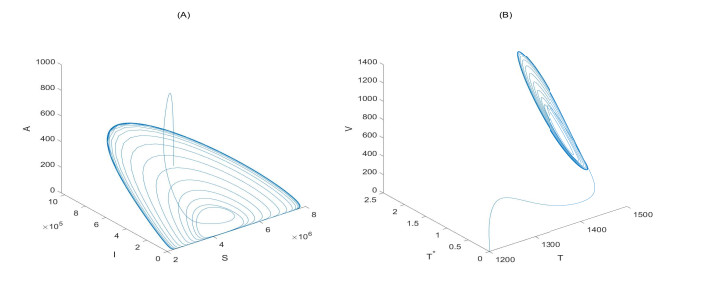
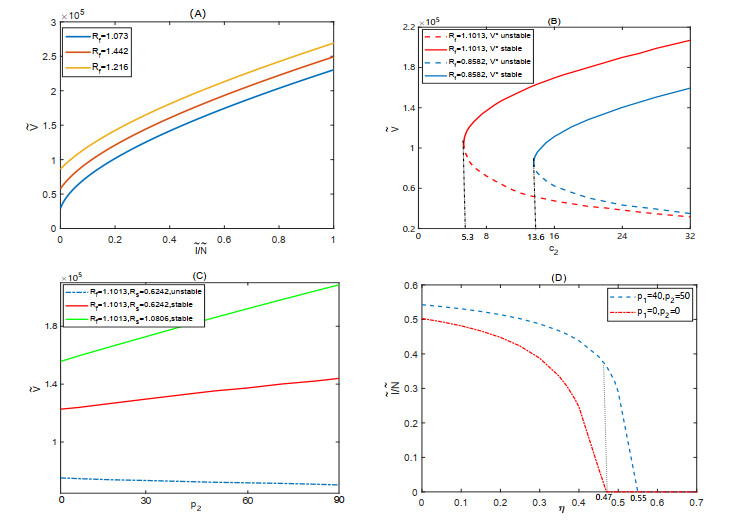
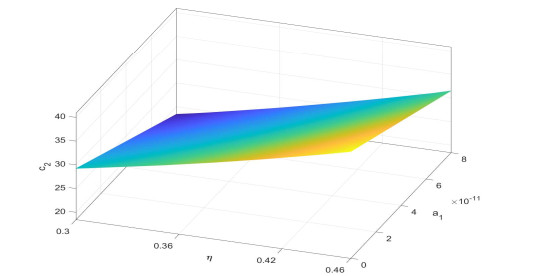
Figures(6)

Yuyi Xue, Yanni Xiao. Analysis of a multiscale HIV-1 model coupling within-host viral dynamics and between-host transmission dynamics[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6720-6736. doi: 10.3934/mbe.2020350







 DownLoad:
DownLoad: