Citation: Simone Göttlich, Stephan Knapp, Dylan Weber. The food seeking behavior of slime mold: a macroscopic approach[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6631-6658. doi: 10.3934/mbe.2020345

| [1] |
T. Nakagaki, Smart behavior of true slime mold in a labyrinth, Res. Microbiol., 152 (2001), 767-770. doi: 10.1016/S0923-2508(01)01259-1

|
| [2] |
T. Nakagaki, R. Kobayashi, Y. Nishiura, T.Ueda, Obtaining multiple separate food sources: behavioural intelligence in the Physarum plasmodium, Proc. R. Soc. London, Ser. B, 271 (2004), 2305-2310. doi: 10.1098/rspb.2004.2856
|
| [3] |
A. Adamatzky, From reaction-diffusion to Physarum computing, Nat. Comput., 8 (2009), 431- 447. doi: 10.1007/s11047-009-9120-5
|
| [4] |
A. Adamatzky, Developing proximity graphs by P. polycephalum: Does the plasmodium follow the toussaint hierarchy?, Parallel Process. Lett., 19 (2009), 105-127. doi: 10.1142/S0129626409000109
|
| [5] |
W. Baumgarten, T. Ueda, M. J. B. Hauser, Plasmodial vein networks of the slime mold Physarum polycephalum form regular graphs, Phys. Rev. E, 82 (2010), 046113. doi: 10.1103/PhysRevE.82.046113
|
| [6] |
T. Latty, M. Beekman, Speed-accuracy trade-offs during foraging decisions in the acellular slime mould Physarum polycephalum, Proc. R. Soc. B, 278 (2011), 539-545. doi: 10.1098/rspb.2010.1624
|
| [7] |
V. Bonifaci, K. Mehlhorn, G. Varma, Physarum can compute shortest paths, J. Theor. Biol., 309 (2012), 121-133. doi: 10.1016/j.jtbi.2012.06.017
|
| [8] | A. Adamatzky, Slime mold solves maze in one pass, assisted by gradient of chemo-attractants, IEEE Trans. NanoBioscience, 11 (2012), 131-134. |
| [9] | C. Oettmeier, K. Brix, H. G. D?bereiner, Physarum polycephalum-a new take on a classic model system, J. Phys. D, 50 (2017), 413001. |
| [10] |
A. Tero, R. Kobayashi, T. Nakagaki, Physarum solver: A biologically inspired method of roadnetwork navigation, Phys. A, 363 (2006), 115-119. doi: 10.1016/j.physa.2006.01.053
|
| [11] | J. Jones, In Approximating the Behaviours of Physarum polycephalum for the Construction and Minimisation of Synthetic Transport Networks, International Conference on Unconventional Computation, Berlin, 2009, Springer, Berlin, 2009,191-208. |
| [12] |
J. Jones, Characteristics of pattern formation and evolution in approximations of Physarum transport Networks, Artif. Life, 16 (2010), 127-153. doi: 10.1162/artl.2010.16.2.16202
|
| [13] |
J. Jones, Influences on the formation and evolution of Physarum polycephalum inspired emergent transport networks, Nat. Comput., 10 (2011), 1345-1369. doi: 10.1007/s11047-010-9223-z
|
| [14] | A. Adamatzky, P. P. B. de Oliveira, Brazilian highways from slime mold's point of view, Kybernetes, 40 (2011), 1373-1394. |
| [15] | J. N. Burchett, O. Elek, N. Tejos, J. X. Prochaska, T. M. Tripp, R. Bordoloi, et al., Revealing the dark threads of the cosmic web, Astrophys. J. Lett., 891 (2020), L35. |
| [16] |
Y. P. Gunji, T. Shirakawa, T. Niizato, T. Haruna, Minimal model of a cell connecting amoebic motion and adaptive transport networks, J. Theor. Biol., 253 (2008), 659-667. doi: 10.1016/j.jtbi.2008.04.017
|
| [17] | M. A. I. Tsompanas, G. Ch Sirakoulis, Modeling and hardware implementation of an amoeba-like cellular automaton, Bioinspiration Biomimetics, 7 (2012), 036013. |
| [18] | Y. X. Liu, Z. L. Zhang, C. Gao, Y. H. Wu, Q. Tao, In A Physarum network evolution model based on IBTM, Advances in swarm intelligence, lecture notes in Computer Science, Berlin, 2013, Y. Tan, Y. H. Shi, H. W. Mo, Eds, Springer, Berlin, 2013, 19-26. |
| [19] |
Y. H. Wu, Z. L. Zhang, Y. Deng, H. Zhou, T. Qian, A new model to imitate the foraging behavior of Physarum polycephalum on a nutrient-poor substrate, Neurocomputing, 148 (2015), 63-69. doi: 10.1016/j.neucom.2012.10.044
|
| [20] | Y. X. Liu, C. Gao, Z. L. Zhang, Y. H. Wu, M. X. Liang, L. Tao, et al., A new multi-agent system to simulate the foraging behaviors of Physarum, Nat. Comput., 16 (2017), 15-29. |
| [21] |
A. Tero, R. Kobayashi, T. Nakagaki, A mathematical model for adaptive transport network in path finding by true slime mold, J. Theor. Biol., 244 (2007), 553-564. doi: 10.1016/j.jtbi.2006.07.015
|
| [22] |
A. Tero, K. Yumiki, R. Kobayashi, T. Saigusa, T. Nakagaki, Flow-network adaptation in Physarum amoebae, Theory Biosci., 127 (2008), 89-94. doi: 10.1007/s12064-008-0037-9
|
| [23] |
R. Borsche, S. Göttlich, A. Klar, P. Schillen, The scalar keller-segel model on networks, Math. Models Methods Appl. Sci., 24 (2014), 221-247. doi: 10.1142/S0218202513400071
|
| [24] | G. Bretti, R. Natalini, Numerical approximation of nonhomogeneous boundary conditions on networks for a hyperbolic system of chemotaxis modeling the Physarum dynamics, J. Comput. Methods Sci. Eng., 18 (2018), 85-115. |
| [25] | J. A. Carrillo, K. Craig, F. S. Patacchini, A blob method for diffusion, Calculus Var. Partial Differ. Equations, 58 (2019), 53. |
| [26] |
L. Chen, S. Göttlich, Q. Yin, Mean Field Limit and Propagation of Chaos for a Pedestrian Flow Model, J. Stat. Phys., 166 (2017), 211-229. doi: 10.1007/s10955-016-1679-5
|
| [27] |
L. Chen, S. Göttlich, S. Knapp, Modeling of a diffusion with aggregation: rigorous derivation and numerical simulation, ESAIM: Math. Modell. Numer. Anal., 52 (2018), 567-593 doi: 10.1051/m2an/2018028
|
| [28] | B. Øksendal, Stochastic Differential Equations, 2nd edition, Springer, Berlin, 2003. |
| [29] | T. Nakagaki, H. Yamada, á. Tóth, Maze-solving by an amoeboid organism, Nature, 407 (2000), 470. |
| [30] | T. Nakagaki, H. Yamada, á. Tóth, Path finding by tube morphogenesis in an amoeboid organism, Biophys. Chem., 92 (2001), 47-52. |
| [31] | L. C. Evans, Partial Differential Equations, 2nd edition, American Mathematical Society, Rhode Island, 2010. |
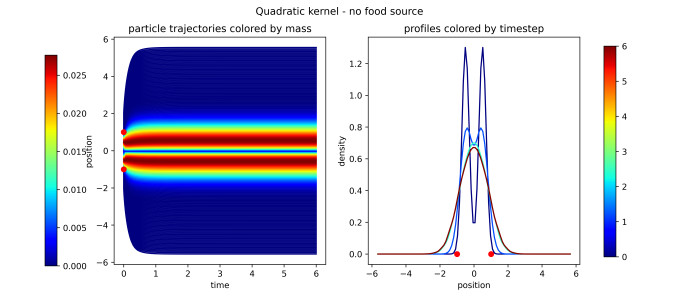
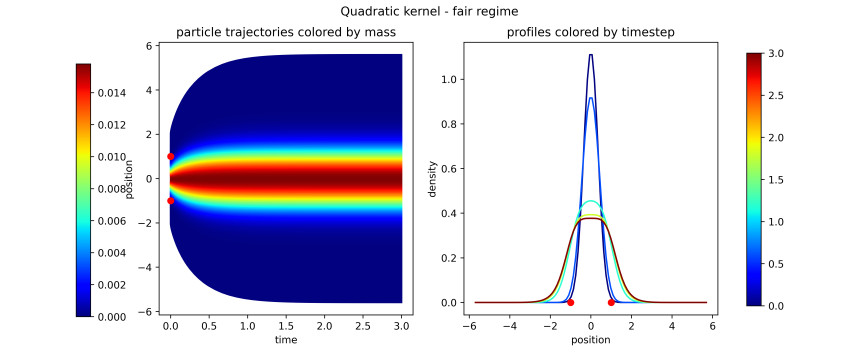
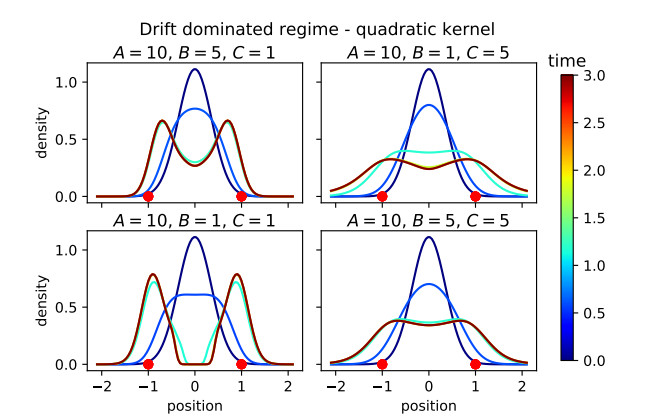
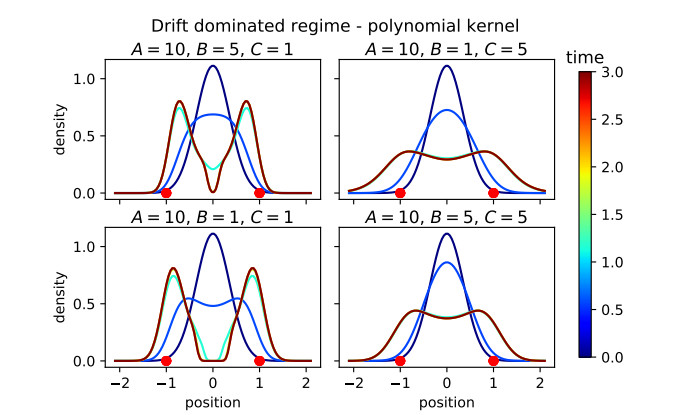
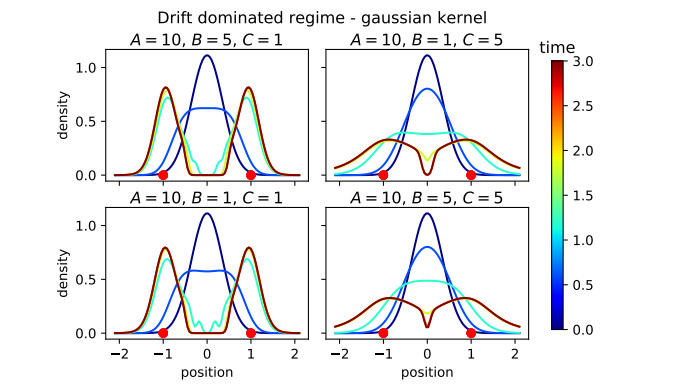
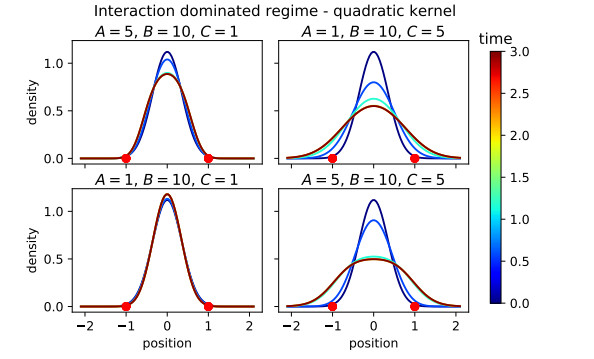
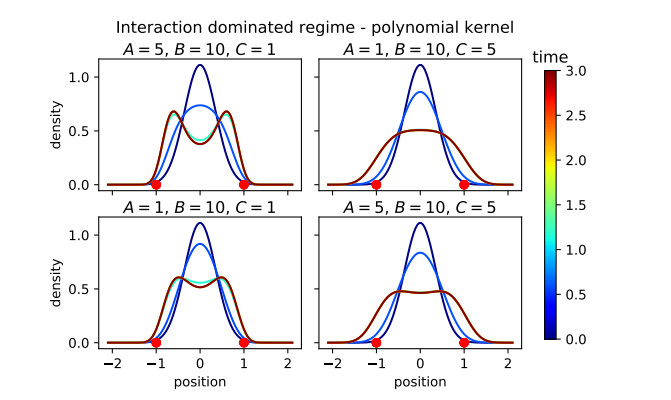
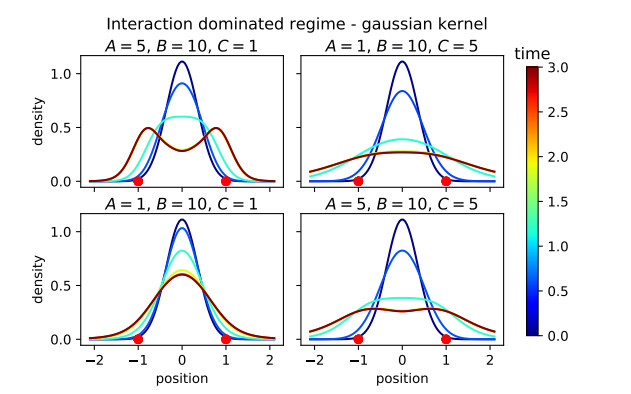
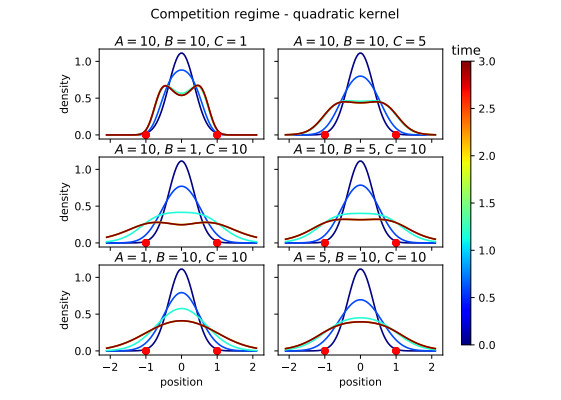
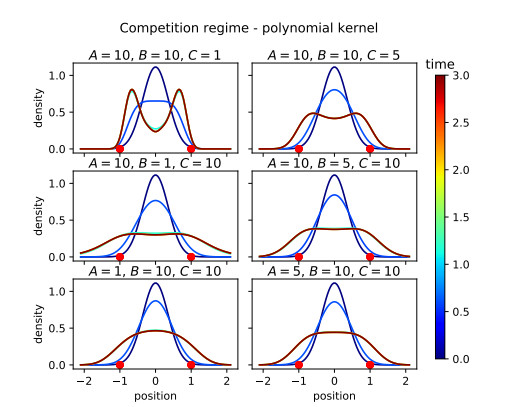
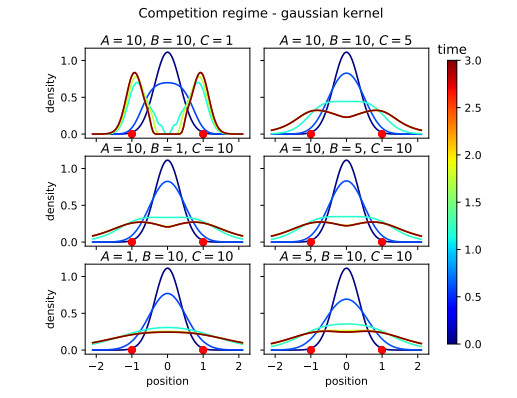
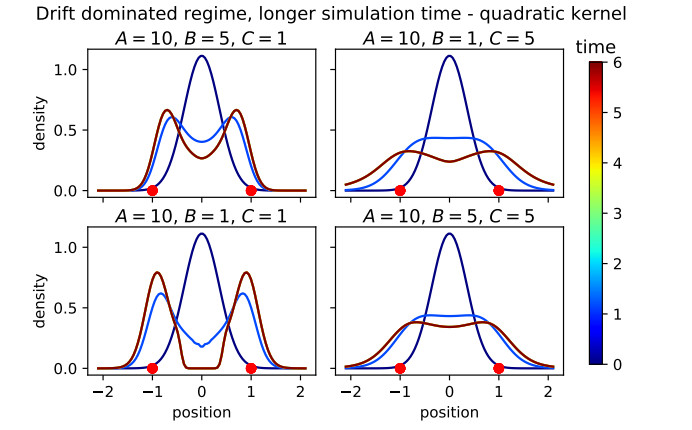
Figures(16)

Simone Göttlich, Stephan Knapp, Dylan Weber. The food seeking behavior of slime mold: a macroscopic approach[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6631-6658. doi: 10.3934/mbe.2020345







 DownLoad:
DownLoad: