In this paper, a reaction-diffusion SI epidemic model with media impact is considered. The boundedness of system and the existence of the state are given. The local stabilities of the endemic states are analyzed. Sufficient conditions of the occurrence of the Turing pattern are obtained by the center manifold theorem and normal form method. Some numerical simulations are given to check in the theoretical results. We find that the influence of media not only inhibits the spread of infectious diseases, but also effects the spatial steady-state of model.
Citation: Xin-You Meng, Tao Zhang. The impact of media on the spatiotemporal pattern dynamics of a reaction-diffusion epidemic model[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4034-4047. doi: 10.3934/mbe.2020223
In this paper, a reaction-diffusion SI epidemic model with media impact is considered. The boundedness of system and the existence of the state are given. The local stabilities of the endemic states are analyzed. Sufficient conditions of the occurrence of the Turing pattern are obtained by the center manifold theorem and normal form method. Some numerical simulations are given to check in the theoretical results. We find that the influence of media not only inhibits the spread of infectious diseases, but also effects the spatial steady-state of model.

| [1] |
A. L. Lloyd, V. A. A. Jansen, Spatiotemporal dynamics of epidemics: Synchrony in metapopulation models, Math. Biosci., 188 (2004), 1-16. doi: 10.1016/j.mbs.2003.09.003

|
| [2] |
M. Su, C. Hui, The effect of predation on the prevalence and aggregation of pathogens in prey, Biosystems, 105 (2011), 300-306. doi: 10.1016/j.biosystems.2011.05.012
|
| [3] | C. Bret, Modeling and inference for infectious disease dynamics: A likelihood-based approach, Stat. Sci., 33 (2018), 57-69. |
| [4] |
G. Q. Sun, J. H. Xie, S. H. Huang, Z. Jin, M.-T. Li, L. Liu, Transmission dynamics of cholera: Mathematical modeling and control strategies, Commun. Nonlin. Sci. Numer. Si., 45 (2017), 235-244. doi: 10.1016/j.cnsns.2016.10.007
|
| [5] |
Y. Kazuo, Threshold dynamics of reaction-diffusion partial differential equations model of Ebola virus disease, Int. J. Biomath., 11 (2018), 1850108. doi: 10.1142/S1793524518501085
|
| [6] | J. R. Petrella, W. R. Hao, A. Rao, P. M. Doraiswamy, Computational causal modeling of the dynamic biomarker cascade in alzheimer's disease, Comput. Math. Methods Med., 2019 (2019), 6216530. |
| [7] |
Q. Yang, H. F. Huo, Dynamics of an edge-based SEIR model for sexually transmitted diseases, Math. Biosci. Eng., 17 (2020), 669-699. doi: 10.3934/mbe.2020035
|
| [8] |
D. M. Xiao, S. G. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Math. Biosci., 208 (2007), 419-429. doi: 10.1016/j.mbs.2006.09.025
|
| [9] | M. Haque, J. Zhen, E. Venturino, An ecoepidemiological predator-prey model with standard disease incidence, Math. Method Appl. Sci., 32 (2010), 875-898. |
| [10] |
L. I. Bo, S. L. Yuan, W. G. Zhang, Analysis on an epidemic model with a ratio-dependent nonlinear incidence rate, Int. J. Biomath., 4 (2011), 227-239. doi: 10.1142/S1793524511001374
|
| [11] | Y. L. Cai, W. M. Wang, Spatiotemporal dynamics of a reaction-diffusion epidemic model with nonlinear incidence rate, J. Stat. Mech. Theory. E, 2011(2011), P02025. |
| [12] |
X. B. Zhang, S. Q. Chang, H. Huo, Dynamic behavior of a stochastic SIR epidemic model with vertical transmission, Electron. J. Differ. Equat., 2019(2019), 1-20. doi: 10.1186/s13662-018-1939-6
|
| [13] | D. Wodarz, D. N. Levy, N. L. Komarova, Multiple infection of cells changes the dynamics of basic viral evolutionary processes, Evolution, 3 (2019), 104-115. |
| [14] |
D. X. Jia, T. H. Zhang, S. L. Yuan, Pattern dynamics of a diffusive toxin producing phytoplanktonzooplankton model with three-dimensional patch, Int. J. Bifurcat. Chaos, 29 (2019), 1930011. doi: 10.1142/S0218127419300118
|
| [15] |
M. Bendahmane, M. Langlais, A reaction-diffusion system with cross-diffusion modeling the spread of an epidemic disease, J. Evol. Equ., 10 (2010), 883-904. doi: 10.1007/s00028-010-0074-y
|
| [16] | W. Z. Huang, M. A. Han, K. Y. Liu, Dynamics of an SIS reaction-diffusion epidemic model for disease transmission, Math. Biosci. Eng., 7 (2017), 51-66. |
| [17] |
S. N. Zhao, S. L. Yuan, H. Wang, Threshold behavior in a stochastic algal growth model with stoichiometric constraints and seasonal variation, J. Differ. Equat., 268 (2020), 5113-5139. doi: 10.1016/j.jde.2019.11.004
|
| [18] | A. D'Onofrio, Stability properties of pulse vaccination strategy in SEIR epidemic model, Math. Biosci., 179(2002), 57-72. |
| [19] | M. DelaSen, S. Alonso-Quesada, R. Nistal, A. Ibeas, On the existence of equilibrium points, boundedness, oscillating behavior and positivity of a sveirs epidemic model under constant and impulsive vaccination, Advances Differ. Equat., 2011 (2011), 339-370. |
| [20] |
A. L. Shane, R. K. Mody, J. A. Crump, L. K. Pickering, 2017 infectious diseases society of America clinical practice guidelines for the diagnosis and management of infectious diarrhea, Clin. Infect. Dis., 65(2017), 1963-1973. doi: 10.1093/cid/cix959
|
| [21] |
C. Kuttler, Modeling to inform infectious disease control, Biometrics, 74 (2018), 382-383. doi: 10.1111/biom.12852
|
| [22] | S. K. Nandi, S. Jana, M. Mandal, K. K. Tapan, Complex dynamics and optimal treatment of an epidemic model with two infectious diseases, Biosystems, 5 (2019), 5-29. |
| [23] |
C. Y. Zheng, C. Y. Xia, Q. T. Guo, M. Dehmer, Interplay between SIR-based disease spreading and awareness diffusion on multiplex networks, J. Parallel Distr. Com., 115 (2018), 20-28. doi: 10.1016/j.jpdc.2018.01.001
|
| [24] |
L. J. S. Allen, B. M. Bolker, Y. Lou, A. L. Nevai, Asymptotic profiles of the steady states for an SIS epidemic patch model, SIAM. J. Appl. Math., 67 (2007), 1283-1309. doi: 10.1137/060672522
|
| [25] |
R. H. Cui, Y. Lou, A spatial SIS model in advective heterogeneous environments, J. Differ. Equat., 261 (2016), 3305-3343. doi: 10.1016/j.jde.2016.05.025
|
| [26] | G. Q. Sun, J. Zhang, L. P. Song, Z. Jin, B. L. Li, Pattern formation of a spatial predator-prey system, Appl. Math. Comput., 218 (2012), 11151-11162. |
| [27] |
G. Q. Sun, L. Li, Z. K. Zhang, Spatial dynamics of a vegetation model in an arid flat environment, Nonlin. Dynam., 73 (2013), 2207-2219. doi: 10.1007/s11071-013-0935-3
|
| [28] | D. Kalajdzievska, M. Y. Li, Modeling the effects of carriers on transmission dynamics of infectious diseases, Math. Biosci. Eng., 8 (2017), 711-722. |
| [29] | F. Berezovskaya, G. Karev, B. J. Song, C. Castillo-Chvez, A simple epidemic model with surprising dynamics, Math. Biosci. Eng., 2 (2005), 133-152. |
| [30] | X. W. Yu, S. L. Yuan, Asymptotic properties of a stochastic chemostat model with two distributed delays and nonlinear perturbation, Discrete Cont. Dyn. Syst. B, 25 (2020), 2373-2390. |
| [31] |
X. Y. Meng, H. F. Huo, X. B. Zhang, Stability and global Hopf bifurcation in a Leslie-Gower predator-prey model with stage structure for prey, J. Appl. Math. Comput., 60 (2019), 1-25. doi: 10.1007/s12190-018-1201-0
|
| [32] |
X. Y. Meng, J. Li, Stability and Hopf bifurcation analysis of a delayed phytoplankton-zooplankton model with Allee effect and linear harvesting, Math. Biosci. Eng., 17 (2020), 1973-2002. doi: 10.3934/mbe.2020105
|
| [33] | X. Y. Meng, Y. Q. Wu, Dynamical analysis of a fuzzy phytoplankton-zooplankton model with refuge, fishery protection and harvesting, J. Appl. Math. Comput., 63 (2020), 361-389. |
| [34] |
W. M. Wang, Y. L. Cai, M. J. Wu, K. F. Wang, Z. Q. Li, Complex dynamics of a reaction-diffusion epidemic model, Nonlin. Anal. Real Word Appl., 13 (2012), 2240-2258. doi: 10.1016/j.nonrwa.2012.01.018
|
| [35] |
T. Wang, Dynamics of an epidemic model with spatial diffusion, Phys. A, 409 (2014), 119-129. doi: 10.1016/j.physa.2014.04.028
|
| [36] |
W. M. Wang, H. Y. Liu, Y. L. Cai, Z. Q. Li, Turing pattern selection in a reaction-diffusion epidemic model, Chinese Phys. B, 20 (2011), 074702. doi: 10.1016/j.physa.2014.04.028
|
| [37] |
M. Fras, M. Gosak, Spatiotemporal patterns provoked by environmental variability in a predatorprey model, Biosystems, 114 (2013), 172-177. doi: 10.1016/j.biosystems.2013.09.004
|
| [38] |
Z. P. Ma, H. F. Huo, H. Xiang, Spatiotemporal patterns induced by delay and cross fractional diffusion in a predator-prey model describing intraguild predation, Math. Methods Appl. Sci., 43 (2020), 5179-5196. doi: 10.1002/mma.6259
|
| [39] |
A. Onofrio, M. Banerjee, P. Manfredi, Spatial behavioural responses to the spread of an infectious disease can suppress Turing and Turing-Hopf patterning of the disease, Phys. A, 545 (2020), 123773. doi: 10.1016/j.physa.2019.123773
|
| [40] | Y. F. Li, J. A. Cui, The effect of constant and pulse vaccination on SIS epidemic models incorporating media coverage, Commun. Nonlin. Sci. Numer. Simul., 14 (2008), 2353-2365. |
| [41] |
M. J. Ma, S. Y. Liu, J. Li, Does media coverage influence the spread of drug addiction, Commun. Nonlinear Sci. Numer. Simul., 50(2017), 169-179. doi: 10.1016/j.cnsns.2017.03.002
|
| [42] | M. A. Khan, S. Islam, G. Zaman, Media coverage campaign in hepatitis b transmission model, Appl. Math. Comput., 331 (2018), 378-393. |
| [43] |
R. S. Liu, J. H. Wu, H. P. Zhu, Media/psychological impact on multiple outbreaks of emerging infectious diseases, Comput. Math. Methods Med., 8 (2007), 153-164. doi: 10.1080/17486700701425870
|
| [44] | J. A. Cui, X. Tao, H. P. Zhu, An SIS infection model incorporating media coverage, Rocky Mountauin J. Math., 38 (2008), 1323-1334. |
| [45] |
Y. P. Liu, J. A. Cui, The impact of media coverage on the dynamics of infections disease, Int. J. Biomath, 1 (2008), 65-74. doi: 10.1142/S1793524508000023
|
| [46] |
Y. F. Li, J. A. Cui, The effect of constant and pulse vaccination on SIS epidemic models incorporating media coverage, Commun. Nonlin. Sci. Numer. Simul., 14 (2009), 2353-2365. doi: 10.1016/j.cnsns.2008.06.024
|
| [47] |
J. A. Cui, Y. H. Sun, H. P. Zhu, The impact of media on the control of infectious diseases, J. Dyn. Differ. Equ., 20 (2008), 31-53. doi: 10.1007/s10884-007-9075-0
|
| [48] |
Y. N. Xiao, S. Y. Tang, J. H. Wu, Media impact switching surface during an infectious disease outbreak, Sci. Rep., 5 (2015), 7838. doi: 10.1038/srep07838
|
| [49] |
Q. L. Yan, S. Y. Tang, S. Gabriele, J. H. Wu, Media coverage and hospital notifications: Correlation analysis and optimal media impact duration to manage a pandemic, J. Theor. Biol., 390 (2016), 1-13. doi: 10.1016/j.jtbi.2015.11.002
|
| [50] | J. M. Tchuenche, N. Dube, B. P. Claver, J. S. Robert, T. B. Chris, The impact of media coverage on the transmission dynamics of human influenza, BMC Public Health, 11(2011), S5. |
| [51] |
C. J. Sun, W. Yang, J. Arino, K. Kamran, Effect of media-induced social distancing on disease transmission in a two patch setting, Math. Biosci., 230 (2011), 87-95. doi: 10.1016/j.mbs.2011.01.005
|
| [52] |
P. V. D. Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1364/JOSA.48.000784
|
| [53] |
X. Y. Meng, J. G. Wang, Analysis of a delayed diffusive model with Beddington-Deangelis functional response, Int. J. Biomath., 12 (2019), 1950047. doi: 10.1142/S1793524519500475
|
| [54] | Y. Zhao, J. Li, X. Ma, Stochastic periodic solution of a susceptible-infective epidemic model in a polluted environment under environmental fluctuation, Comput. Math. Methods Med., 2018 (2018), 1-15. |
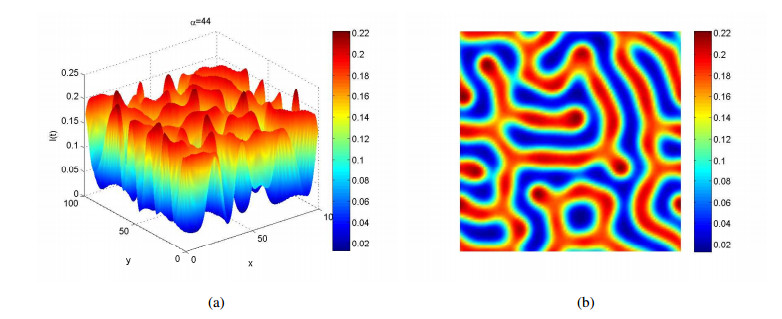
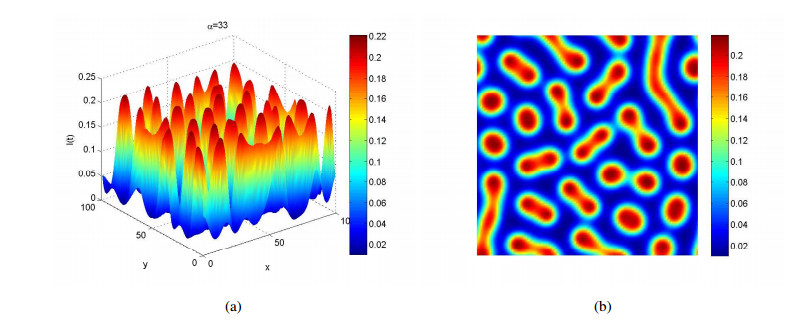
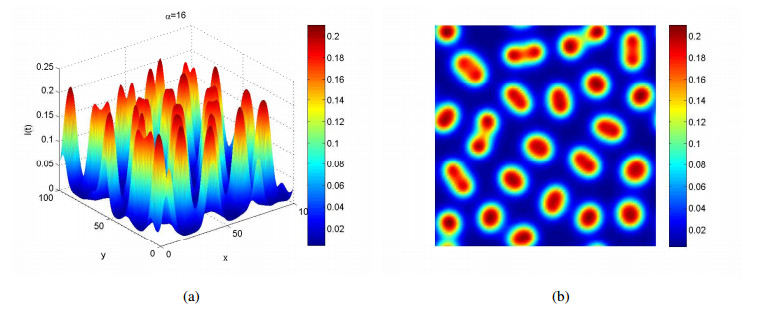
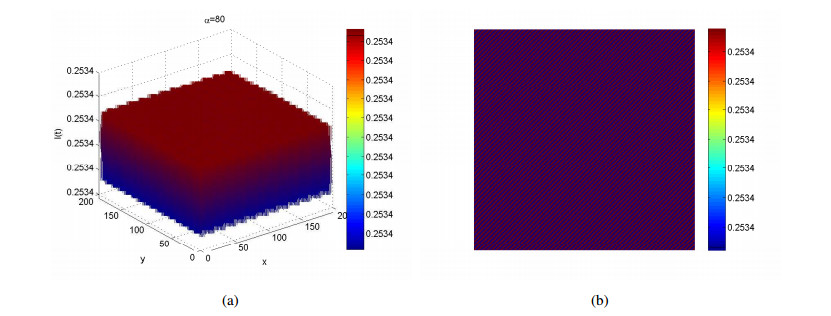
Figures(4)

Xin-You Meng, Tao Zhang. The impact of media on the spatiotemporal pattern dynamics of a reaction-diffusion epidemic model[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 4034-4047. doi: 10.3934/mbe.2020223







 DownLoad:
DownLoad: