Citation: Qian Ding, Jian Liu, Zhiming Guo. Dynamics of a malaria infection model with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4885-4907. doi: 10.3934/mbe.2019246

| [1] | WHO, "Malaria", 2008. Available from: http://www.who.int/malaria/en. |
| [2] | F. McKenzie and H. Bossert, An integrated model of Plasmodium falciparum dynamics, J. Theoret. Biol., 232 (2005), 411–426. |
| [3] | P. Streatfield, W. Khan and A. Bhuiya, Malaria mortality in Africa and Asia: evidence from INDEPTH health and demographic surveillance model sites, Global Health Action, 7 (2014), 25369–25369. |
| [4] | C. Hetzel and R. Anderson, The within-host cellular dynamics of bloodstage malaria: theoretical and experimental studies, Parasitology, 113 (1996), 25–38. |
| [5] | Y. Li, S. Ruan and D. Xiao, The within-host dynamics of malaria infection with immune response, Math. Biosci. Eng., 8 (2011), 999–1018. |
| [6] | Y. Xiao and X. Zou, Can Multiple Malaria Species Co-persist?, SIAM J. Appl. Math., 73 (2013), 351–373. |
| [7] | Z. Chang, J. Ning and Y. Zhang, The TatD-like DNase of Plasmodium is a virulence factor and a potential malaria vaccine candidate, Nat. Commun., 7 (2016), 11537–11546. |
| [8] | L. Molineaux and K. Dietz, Review of intra-host models of malaria, Parassitologia, 41 (1999), 221–231. |
| [9] | R. Anderson, R. May and S. Gupta, Non-linear phenomena in host-parasite interactions, Parasitology, 99 (1989), S59–S79. |
| [10] | C. Chiyaka, W. Garira and S. Dube, Modelling immune response and drug therapy in human malaria infection, Comput. Math. Methods Med., 9 (2008), 143–163. |
| [11] | G. Johnston, P. Gething and S. Hay, Modeling within-host effects of drugs on Plasmodium falciparum transmission and prospects for malaria elimination, PLoS Comput. Biol., 10 (2014), e1003434. |
| [12] | K. Landman, K. Tan and P. Arguin, Adherence to malaria prophylaxis among Peace Corps Volunteers in the Africa region, Travel. Med. Infect. Dis., 13 (2015), 61–68. |
| [13] | J. Liu and Z. Guo, The dynamic behavior of mathematical model of pathogenesis of plasmodium falciparum, J. Guangzhou Univ., 17 (2018), 13–22. |
| [14] | Y. Kuang, Delay differential equations with applications in population dynamics, Academic Press, Boston, 1993. |
| [15] | H. L. Smith, An introduction to delay differential equations with applications to the life sciences, Springer, New York, 2011. |
| [16] | J. Hale, Oscillations in neutral functional differential equations, C.i.m.e.summer Schools, 1972, 97–111. |
| [17] | Z. Wang, Z. Guo and H. Peng, A mathematical model verifying potent oncolytic efficacy of M1 virus, Math. Biosci., 276 (2016), 19–27. |
| [18] | S. Ruan and W. Wang, Dynamical behavior of an epidemic model with a nonlinear incidence rate, J. Differential Equations, 188 (2003), 135–163. |
| [19] | D. Xiao and Y. Yang, Influence of latent period and nonlinear incidence rate on the dynamics of SIRS epidemiological models, Discrete Contin. Dyn. Syst. Ser. B, 13 (2012), 195-211. |
| [20] | B. D. Hassard, N. D. Kazarinoff and Y. Wan, Theory and applications of Hopf bifurcation, Cambridge University Press, Cambridge, 1981. |
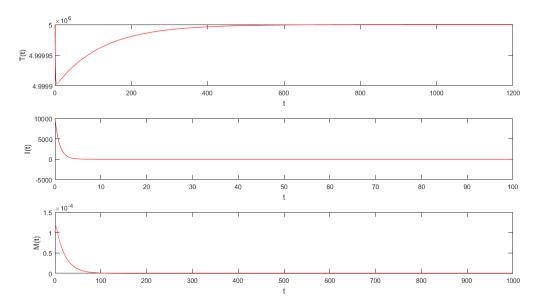
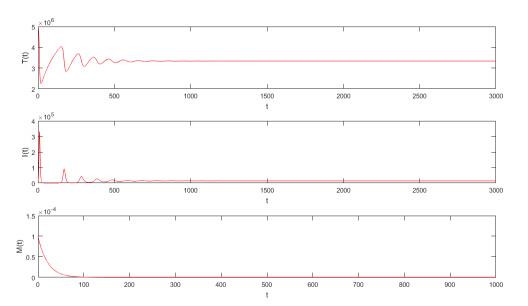
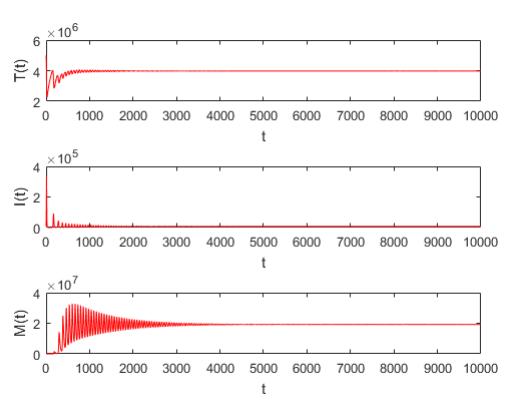
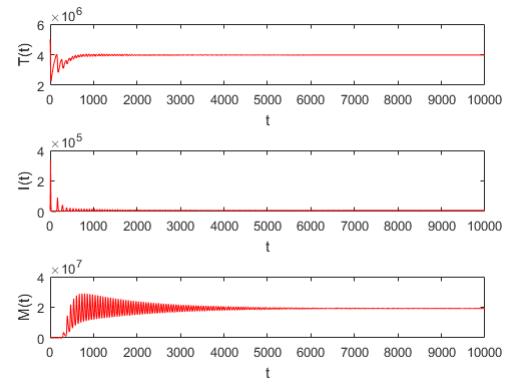
Figures(5) / Tables(2)

Qian Ding, Jian Liu, Zhiming Guo. Dynamics of a malaria infection model with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4885-4907. doi: 10.3934/mbe.2019246







 DownLoad:
DownLoad: