Citation: Peter Rashkov, Ezio Venturino, Maira Aguiar, Nico Stollenwerk, Bob W. Kooi. On the role of vector modeling in a minimalistic epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4314-4338. doi: 10.3934/mbe.2019215

| [1] | N. Ferguson, R. Anderson and S. Gupta, The effect of antibody-dependent enhancement on the transmission dynamics and persistence of multiple-strain pathogens, Proc. Natl. Acad. Sci. USA, 96 (1999), 790–794. |
| [2] | L. Billings, I. B. Schwartz, L. B. Shaw, et al., Instabilities in multiserotype disease models with antibody-dependent enhancement, J. Theor. Biol., 246 (2007), 18–27. |
| [3] | M. Aguiar, S. Ballesteros, B. W. Kooi, et al., The role of seasonality and import in a minimalistic multi-strain dengue model capturing differences between primary and secondary infections: complex dynamics and its implications for data analysis, J. Theor. Biol., 289 (2011), 181–196. |
| [4] | B. W. Kooi, M. Aguiar and N. Stollenwerk, Bifurcation analysis of a family of multi-strain epidemiology models, J. Comput. Appl. Math., 252 (2013), 148–158. |
| [5] | T.-T. Zheng and L.-F. Nie, Modelling the transmission dynamics of two-strain Dengue in the presence awareness and vector control, J. Theor. Biol., 443 (2018), 82–91. |
| [6] | Z. Feng and J. X. Velasco-Hernandez, Competitive exclusion in a vector-host model for the dengue fever, J. Math. Biol., 35 (1997), 523–544. |
| [7] | M. Zhu and Y. Xu, A time-periodic dengue fever model in a heterogeneous environment, Math. Comput. Simulat., 155 (2019), 115–129. |
| [8] | B. Althouse, J. Lessler, A. Sall, et al., Synchrony of sylvatic dengue isolations: A multi-host, multi-vector SIR model of dengue virus transmission in Senegal, PLoS Negl. Trop. Dis., 6 (2012), e1928. |
| [9] | F. Rocha, M. Aguiar, M. Souza, et al., Time-scale separation and centre manifold analysis describing vector-borne disease dynamics, Int. J. Comput. Math., 90 (2013), 2105–2125. |
| [10] | A. Segel and M. Slemrod, The Quasi-Steady-State Assumption: A case study in perturbation, SIAM Rev., 31 (1989), 446–477. |
| [11] | F. Rocha, L. Mateus, U. Skwara, et al., Understanding dengue fever dynamics: a study of seasonality in vector-borne disease models, Int. J. Comput. Math., 93 (2016), 1405–1422. |
| [12] | T. G¨otz, K. P. Wijaya and E. Venturino, Introducing seasonality in an SIR-UV epidemic model: an application to dengue, in 18th International Conference on Computational and Mathematical Methods in Science and Engineering, CMMSE 2018 (ed. J. V. Aguiar), Wiley Online Library, 2018, Online ISSN:2577-7408. |
| [13] | Y. Nagao and K. Koelle, Decreases in dengue transmission may act to increase the incidence of dengue hemorrhagic fever, Proc. Natl. Acad. Sci. USA, 105 (2008), 2238–2243. |
| [14] | C. K. R. T. Jones, Geometric singular perturbation theory, in Dynamical Systems: Lectures Given at the 2nd Session of the Centro Internazionale Matematico Estivo (C.I.M.E.) held in Montecatini Terme, Italy, June 13–22, 1994 (ed. R. Johnson), Springer, Berlin, Heidelberg, 1995, 44–118. |
| [15] | G. Hek, Geometric singular perturbation theory in biological practice, J. Math. Biol., 60 (2010), 347–386. |
| [16] | B. W. Kooi and J.-C. Poggiale, Modelling, singular perturbation and bifurcation analyses of bitrophic food chains, Math. Biosci., 301 (2018), 93–110. |
| [17] | C. S. Holling, Some characteristics of simple types of predation and parasitism, Can. Entomol., 91 (1959), 385–398. |
| [18] | M. L. Rosenzweig and R. H. MacArthur, Graphical representation and stability conditions of predator-prey interactions, Am. Nat., 97 (1963), 209–223. |
| [19] | N. Fenichel, Persistence and smoothness of invariant manifolds for flows, Indiana U. Math. J., 21 (1971), 193–226. |
| [20] | N. Fenichel, Geometric singular perturbation theory, J. Differ. Equations, 31 (1979), 53–98. |
| [21] | A. Korobeinikov, Lyapunov functions and global properties for SEIR and SEIS epidemic models, Math. Med. Biol., 21 (2004), 75–83. |
| [22] | M. Kot, Elements of Mathematical Ecology, Cambridge University Press, Cambridge, 2001. |
| [23] | O. Diekmann, J. A. P. Heesterbeek and J. A. J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. |
| [24] | O. Diekmann and J. A. P. Heesterbeek, Mathematical epidemiology of infectious diseases, Wiley series in mathematical and computional biology, Wiley, Chichester, 2000. |
| [25] | V. Capasso and G. Serio, A generalization of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. |
| [26] | K. Dietz, Overall population patterns in the transmission cycle of infectious disease agents, in Population biology of infectious diseases (eds. R. M. Anderson and R. M. May), Springer, Berlin, Heidelberg, 1982, 87–102. |
| [27] | J. Mena-Lorca and H. W. Hethcote, Dynamic models of infectious diseases as regulators of population size, J. Math. Biol., 30 (1992), 693–716. |
| [28] | B. W. Kooi, G. A. K. van Voorn and K. P. Das, Stabilization and complex dynamics in a predatorprey model with predator suffering from an infectious disease, Ecol. Complex., 8 (2011), 113–122. |
| [29] | F. Irwin and H. N. Wright, Some properties of polynomial curves, Ann. Math. Second Series, 19 (1917), 152–158. |
| [30] | L. Edelstein-Keshet, Mathematical Models in Biology, SIAM, Philadelphia, 2005. |
| [31] | E. J. Doedel and B. Oldeman, AUTO 07p: Continuation and Bifurcation software for ordinary differential equations, Technical report, Concordia University, Montreal, Canada, 2009. |
| [32] | A.-M. Helt and E. Harris, S-phase-dependent enhancement of dengue virus 2 replication in mosquito cells, but not in human cells, J. Virology, 79 (2005), 13218–13230. |
| [33] | J. Hily, A. García, A. Moreno, M. Plaza, M.Wilkinson, A. Fereres, A. Fraile and F. García-Arenal, The relationship between host lifespan and pathogen reservoir potential: An analysis in the system Arabidopsis thaliana-Cucumber mosaic virus, PLoS Pathog., 10 (2014), e1004492. |
| [34] | N. M. Nguyen, D. Thi Hue Kien, T. V. Tuan, et al., Host and viral features of human dengue cases shape the population of infected and infectious Aedes aegypti mosquitoes, Proc. Natl. Acad. Sci. USA, 110 (2013), 9072–9077. |
| [35] | J. Yu, Modeling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. |
| [36] | M. R. W. de Valdez, D. Nimmo, J. Betz, et al., Genetic elimination of dengue vector mosquitoes, Proc. Natl. Acad. Sci. USA, 108 (2011), 4772–4775. |
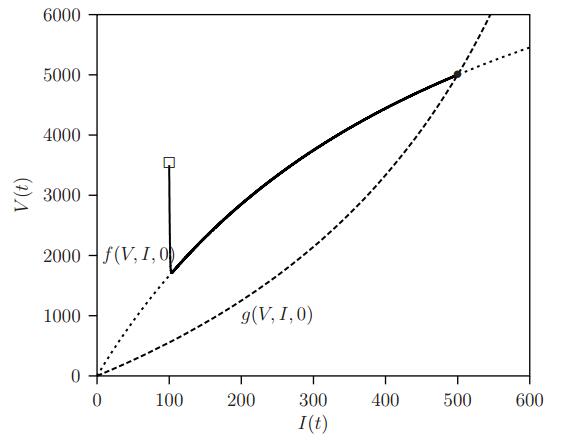
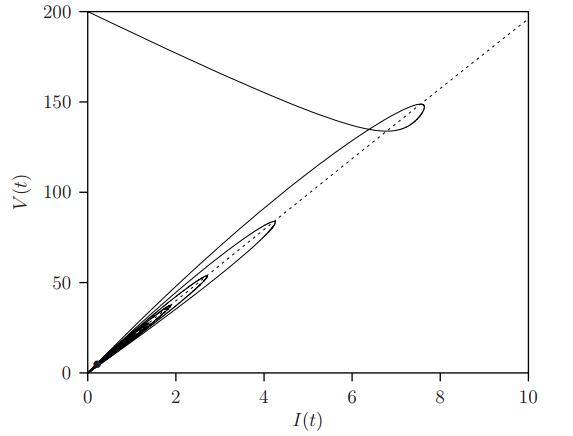
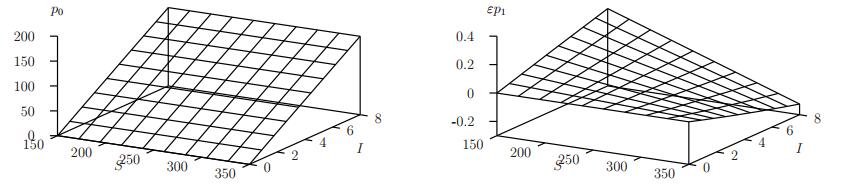

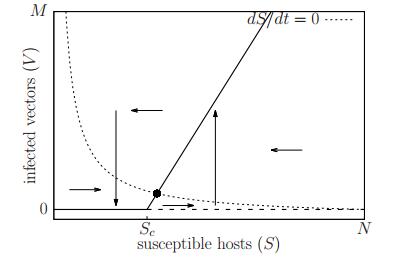
Figures(11) / Tables(1)

Peter Rashkov, Ezio Venturino, Maira Aguiar, Nico Stollenwerk, Bob W. Kooi. On the role of vector modeling in a minimalistic epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4314-4338. doi: 10.3934/mbe.2019215







 DownLoad:
DownLoad: