Citation: Min Zhu, Xiaofei Guo, Zhigui Lin. The risk index for an SIR epidemic model and spatial spreading of the infectious disease[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1565-1583. doi: 10.3934/mbe.2017081

| [1] | [ I. Ahn,S. Baek,Z. G. Lin, The spreading fronts of an infective environment in a man-environment-man epidemic model, Appl. Math. Modelling, 40 (2016): 7082-7101. |
| [2] | [ L. J. S. Allen,B. M. Bolker,Y. Lou,A. L. Nevai, Asymptotic profiles of the steady states for an SIS epidemic reaction-diffusion model, Discrete Contin. Dyn. Syst. Ser. A, 21 (2008): 1-20. |
| [3] | [ A. L. Amadori,J. L. Vázquez, Singular free boundary problem from image processing, Math. Model. Methods. Appl. Sci., 15 (2005): 689-715. |
| [4] | [ A. Bezuglyy,Y. Lou, Reaction-diffusion models with large advection coefficients, Appl. Anal., 89 (2010): 983-1004. |
| [5] | [ R. S. Cantrell and C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, John Wiley & Sons, Ltd, 2003. |
| [6] | [ Center for Disease Control and Prevention (CDC), Update: West Nile-like viral encephalitis-New York, 1999, Morb. Mortal Wkly. Rep., 48 (1999): 890-892. |
| [7] | [ X. F. Chen,A. Friedman, A free boundary problem arising in a model of wound healing, SIAM J. Math. Anal., 32 (2000): 778-800. |
| [8] | [ Y. H. Du,Z. G. Lin, Spreading-vanishing dichotomy in the diffusive logistic model with a free boundary, SIAM J. Math. Anal., 42 (2010): 377-405. |
| [9] | [ Y. H. Du,Z. G. Lin, The diffusive competition model with a free boundary: Invasion of a superior or inferior competitor, Discrete Contin. Dyn. Syst. Ser. B, 19 (2014): 3105-3132. |
| [10] | [ Y. H. Du,B. D. Lou, Spreading and vanishing in nonlinear diffusion problems with free boundaries, J. Eur. Math. Soc., 17 (2015): 2673-2724. |
| [11] | [ A. M. Elaiw,N. H. AlShamrani, Global stability of humoral immunity virus dynamics models with nonlinear infection rate and removal, Nonlinear Anal. Real World Appl., 26 (2015): 161-190. |
| [12] | [ X. M. Feng,S. G. Ruan,Z. D. Teng,K. Wang, Stability and backward bifurcation in a malaria transmission model with applications to the control of malaria in China, Math. Biosci., 266 (2015): 52-64. |
| [13] | [ D. Z. Gao, Y. J. Lou and D. H. He, et al., Prevention and control of Zika as a mosquito-borne and sexually transmitted disease: A mathematical modeling analysis, Scientific Reports 6, Article number: 6 (2016), 28070. |
| [14] | [ J. Ge,K. I. Kim,Z. G. Lin,H. P. Zhu, A SIS reaction-diffusion model in a low-risk and high-risk domain, J. Differential Equation, 259 (2015): 5486-5509. |
| [15] | [ H. Gu,B. D. Lou,M. L. Zhou, Long time behavior of solutions of Fisher-KPP equation with advection and free boundaries, J. Funct. Anal., 269 (2015): 1714-1768. |
| [16] | [ Globalization and disease Available from: https://en.wikipedia.org/wiki/Globalization_and_disease. |
| [17] | [ West African Ebola virus epidemic Available from: https://en.wikipedia.org/wiki/West_African_Ebola_virus_epidemic. |
| [18] | [ Epidemic situation of dengue fever in Guangdong, 2014 (Chinese) Available from: http://www.rdsj5.com/guonei/1369.html. |
| [19] | [ S. Iwami,Y. Takeuchi,X. N. Liu, Avian-human influenza epidemic model, Math. Biosci., 207 (2007): 1-25. |
| [20] | [ W. O. Kermack,A. G. Mckendrick, Contributions to the mathematical theory of epidemics, Proc. Roy. Soc., A115 (1927): 700-721. |
| [21] | [ W. O. Kermack,A. G. Mckendrick, Contributions to the mathematical theory of epidemics, Proc. Roy. Soc., A138 (1932): 55-83. |
| [22] | [ K. I. Kim,Z. G. Lin,L. Zhang, Avian-human influenza epidemic model with diffusion, Nonlinear Anal. Real World Appl., 11 (2010): 313-322. |
| [23] | [ K. I. Kim,Z. G. Lin,Q. Y. Zhang, An SIR epidemic model with free boundary, Nonlinear Anal. Real World Appl., 14 (2013): 1992-2001. |
| [24] | [ O. A. Ladyzenskaja, V. A. Solonnikov and N. N. Ural'ceva, Linear and Quasilinear Equations of Parabolic Type, Amer. Math. Soc, Providence, RI, 1968. |
| [25] | [ C. X. Lei,Z. G. Lin,Q. Y. Zhang, The spreading front of invasive species in favorable habitat or unfavorable habitat, J. Differential Equations, 257 (2014): 145-166. |
| [26] | [ C. Z. Li,J. Q. Li,Z. E. Ma,H. P. Zhu, Canard phenomenon for an SIS epidemic model with nonlinear incidence, J. Math. Anal. Appl., 420 (2014): 987-1004. |
| [27] | [ M. Li,Z. G. Lin, The spreading fronts in a mutualistic model with advection, Discrete Contin. Dyn. Syst. Ser. B, 20 (2015): 2089-2105. |
| [28] | [ Z. G. Lin, A free boundary problem for a predator-prey model, Nonlinearity, 20 (2007): 1883-1892. |
| [29] | [ Z. G. Lin and H. P. Zhu, Spatial spreading model and dynamics of West Nile virus in birds and mosquitoes with free boundary, J. Math. Biol., (2017), 1–29, http://link.springer.com/article/10.1007/s00285-017-1124-7?wt_mc=Internal.Event.1.SEM.ArticleAuthorOnlineFirst. |
| [30] | [ N. A. Maidana,H. Yang, Spatial spreading of West Nile Virus described by traveling waves, J. Theoret. Biol., 258 (2009): 403-417. |
| [31] | [ W. Merz,P. Rybka, A free boundary problem describing reaction-diffusion problems in chemical vapor infiltration of pyrolytic carbon, J. Math. Anal. Appl., 292 (2004): 571-588. |
| [32] | [ L. I. Rubinstein, The Stefan Problem, Amer. Math. Soc, Providence, RI, 1971. |
| [33] | [ C. Shekhar, Deadly dengue: New vaccines promise to tackle this escalating global menace, Chem. Biol., 14 (2007): 871-872. |
| [34] | [ S. Side,S. M. Noorani, A SIR model for spread of Dengue fever disease (simulation for South Sulawesi, Indonesia and Selangor, Malaysia), World Journal of Modelling and Simulation, 9 (2013): 96-105. |
| [35] | [ Y. S. Tao,M. J. Chen, An elliptic-hyperbolic free boundary problem modelling cancer therapy, Nonlinearity, 19 (2006): 419-440. |
| [36] | [ J. Wang,J. F. Cao, The spreading frontiers in partially degenerate reaction-diffusion systems, Nonlinear Anal., 122 (2015): 215-238. |
| [37] | [ J. Y. Wang,Y. N. Xiao,Z. H. Peng, Modelling seasonal HFMD infections with the effects of contaminated environments in mainland China, Appl. Math. Comput., 274 (2016): 615-627. |
| [38] | [ M. X. Wang,J. F. Zhao, Free boundary problems for a Lotka-Volterra competition system, J. Dynam. Differential Equations, 26 (2014): 655-672. |
| [39] | [ World Health Organization, World Health Statistics 2006-2012. |
| [40] | [ P. Zhou,D. M. Xiao, The diffusive logistic model with a free boundary in heterogeneous environment, J. Differential Equations, 256 (2014): 1927-1954. |
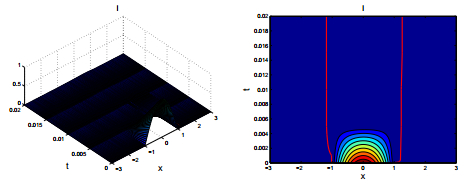
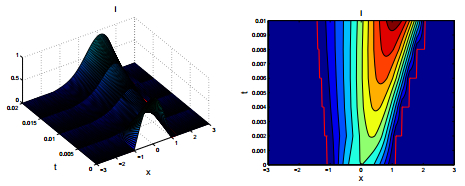
Figures(2)

Min Zhu, Xiaofei Guo, Zhigui Lin. The risk index for an SIR epidemic model and spatial spreading of the infectious disease[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1565-1583. doi: 10.3934/mbe.2017081







 DownLoad:
DownLoad: