Citation: Jorge Duarte, Cristina Januário, Nuno Martins. A chaotic bursting-spiking transition in a pancreatic beta-cells system: observation of an interior glucose-induced crisis[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 821-842. doi: 10.3934/mbe.2017045

| [1] | [ J. Aguirre, E. Mosekilde and M. A. F. Sanjuán, Analysis of the noise-induced bursting-spiking transition in a pancreatic $β $-cell model ,Pysical Review E, 69 (2004), 041910, 16pp. |
| [2] | [ I. Atwater, C. M. Dawson, A. Scott, G. Eddlestone and E. Rojas, The Nature of the Oscillatory Behaviour in Electrical Activity from Pancreatic Beta-cell ,Georg Thieme, New York, 1980. |
| [3] | [ C. A. S. Batista, A. M. Batista, J. A. C. de Pontes, R. L. Viana and S. R. Lopes, Chaotic phase synchronization in scale-free networks of bursting neurons,Phys. Rev. E, 76 (2007), 016218, 10pp. |
| [4] | [ C. A. S. Batista, E. L. Lameu, A. M. Batista, S. R. Lopes, T. Pereira, G. Zamora-López, J. Kurths and R. L. Viana. Phase synchronization of bursting neurons in clustered small-world networks ,Phys. Rev. E ,86 (2012), 016211. |
| [5] | [ R. Bertram,A. Sherman, Dynamical complexity and temporal plasticity in pancreatic beta cells, J. Biosci., 25 (2000): 197-209. |
| [6] | [ T. R. Chay,J. Keizer, Minimal model for membrane oscillations in the pancreatic beta-cel, Biophys. J., 42 (1983): 181-190. |
| [7] | [ T. R. Chay, Chaos in a three-variable model of an excitable cell, Physica D, 16 (1984): 233-242. |
| [8] | [ L. O. Chua,M. Komuro,T. Matsumoto, The double scroll family, IEEE Trans. Circuits Syst., 32 (1985): 797-818. |
| [9] | [ B. Deng, A mathematical model that mimics the bursting oscillations in pancreatic $β $-cells, Math. Biosciences, 119 (1994): 241-250. |
| [10] | [ B. Deng, Glucose-induced period-doubling cascade in the electrical activity of pancreatic $β $-cells, J. Math. Biol., 38 (1999): 21-78. |
| [11] | [ J. Duarte,C. Januário,N. Martins, Topological entropy and the controlled effect of glucose in the electrical activity of pancreatic beta-cells, Physica D, 238 (2009): 2129-2137. |
| [12] | [ J. Duarte,C. Januário,N. Martins, Explicit series solution for a glucose-induced electrical activity model of pancreatic cells, Chaos, Solitons & Fractals, 76 (2015): 1-9. |
| [13] | [ J. Duarte, C. Januário, C. Rodrigues and J. Sardany és, Topological complexity and predictability in the dynamics of a tumor growth model with Shilnikov's chaos ,Int. J Bifurcation Chaos, 23 (2013), 1350124, 12pp. |
| [14] | [ H. Fallah, Symmetric fold / super-Hopf bursting, chaos and mixed-mode oscillations in Pernarowski model Int.,J Bifurcation Chaos, 26 (2016), 1630022, 14pp. |
| [15] | [ Y.-S. Fan,T. R. Chay, Crisis Transitions in excitable cell models, Chaos, Solitons & Fractals, 3 (1993): 603-615. |
| [16] | [ Y.-S. Fan,T. R. Chay, Crisis and topological entropy, Physical Review E, 51 (1995): 1012-1019. |
| [17] | [ L. E. Fridlyand,N. Tamarina,L. H. Philipson, Bursting and calcium oscillations in pancreatic beta cells: Specific pacemakers, Am J Physiol Endocrinol Metab., 299 (2010): E517-E532. |
| [18] | [ J. M. González-Miranda, Observation of a continuous interior crisis in the Hindmarsh-Rose neuron model, Chaos, 13 (2003), 845. |
| [19] | [ J. M. González-Miranda, Complex bifurcations structures in the Hindmarsh-Rose neuron model, Int. J Bifurcation Chaos, 17 (2007): 3071-3083. |
| [20] | [ J. M. González-Miranda, Nonlinear dynamics of the membrane potential of a bursting pacemaker cell, Chaos, 22 (2012), 013123. |
| [21] | [ J. M. González-Miranda, Pacemaker dynamics in the full Morris-Lecar model, Commun. Nonlinear Sci Numer Simulat, 19 (2014): 3229-3241. |
| [22] | [ H.-G. Gu,B. Jia,G.-R. Chen, Experimental evidence of a chaotic region in a neural pacemaker, Physics Letters A, 377 (2013): 718-720. |
| [23] | [ H. Gu,B. Pan,G. Chen, Biological experimental demonstration of bifurcations from bursting to spiking predicted by theoretical models, Nonlinear Dyn., 78 (2014): 391-407. |
| [24] | [ S. Jalil, I. Belykh and A. Shilnikov. Spikes matter for phase-locked bursting in inhibitory neurons ,Phys. Rev. E,85 (2012), 036214. |
| [25] | [ A. Katok and B. Hasselblatt, Introduction to the Modern Theory of Dynamical Systems ,Cambridge University Press, 1995. |
| [26] | [ J. P. Lampreia,J. S. Ramos, Symbolic dynamics of bimodal maps, Portugal. Math., 54 (1997): 1-18. |
| [27] | [ S. J. Liao, Beyond Perturbation: Introduction to the Homotopy Analysis Method ,CRC Press, Chapman and Hall, Boca Raton, FL, 2004. |
| [28] | [ S. J. Liao,Y. Tan, A general approach to obtain series solutions of nonlinear differential equations, Stud. Appl. Math., 119 (2007): 297-354. |
| [29] | [ S. Liao, Advances in the Homotopy Analysis Method, World Scientific Publishing Co, 2014. |
| [30] | [ A. J. Tan,M. A. Lieberman, null, Regular and Chaotic Dynamics, Springer-Verlag, New York, 1992. |
| [31] | [ A. Markovic,T. L. O. Stozer,M. Gosak,J. Dolensek,M. Marhl, Progressive glucose stimulation of islet beta cells reveals a transition from segregated to integrated modular functional connectivity patterns, Scientific Reports, 5 (2015): 7845. |
| [32] | [ T. Matsumoto,T. L. O. Chua,M. Komuro., The double scroll family, IEEE Trans. Circuits Syst., 32 (1985): 797-818. |
| [33] | [ J. Milnor,W. Thurston, On iterated maps of the interval, Lect. Notes in Math., 1342 (1988): 465-563. |
| [34] | [ M. Misiurewicz,W. Szlenk, Entropy of piecewise monotone mappings, Studia Math., 67 (1980): 45-63. |
| [35] | [ S. E. Newhouse,D. Ruelle,F. Takens, Occurrence of strange Axiom A attractors near quasiperiodic flows on T$^{m}$, $m≥3$, Commun. Math. Phys., 64 (1978): 35-40. |
| [36] | [ E. Ott, Chaos in Dynamical Systems ,Cambridge University Press, Cmabridge, UK, 2002. |
| [37] | [ T. Parker and L. O. Chua, Practical Numerical Algorithms for Chaotic Systems ,Springer-Verlag, 1989. |
| [38] | [ M. G. Pederson,E. Mosekilde,K. S. Polonsky,D. S. Luciani, Complex Patterns of Metabolic and Ca$^{2+}$ entrainment in pancreatic islets by oscillatory glucose, Biophysical Journal, 105 (2013): 29-39. |
| [39] | [ K. Ramasubramanian,M. S. Sriram, A Comparative study of computation of Lyapunov spectra with different algorithms, Physica D, 139 (2000): 72-86. |
| [40] | [ J. Rinzel, null, Ordinary and Partial Differential Equations, Springer, New York, 1985. |
| [41] | [ G. A. Rutter,D. J. Hodson, Minireview: Intraislet regulation of insulin secretion in humans, Molecular Endocrinology, 27 (2013): 1984-1995. |
| [42] | [ A. Sherman, P. Carroll, R. M. Santos and I. Atwater, Glucose Dose Response of Pancreatic $β $-cells: Experimental and Theoretical Results, Transitions in Biological Systems, Eds Pienum, New York, 1990. |
| [43] | [ A. Stozer, M. Gosak, J. Dolensek, M. Perc, M. Marhl and M. S. Rupnik, Functional Connectivity in Islets of Langerhans from Mouse Pancreas Tissue Slices, PLoS Comput. Biol. , 9 (2013), e1002923. |
| [44] | [ A. Stozer, M. Gosak, J. Dolensek, M. Perc, M. Marhl and M. S. Rupnik, Functional connectinity in islets of Langerhans from mouse pancreas tissue slices, PLOS Comp. Biol. ,9 (2013), e1002923. |
| [45] | [ J. Wang,S. Liu,X. Liu, Quantification of synchronization phenomena in two reciprocally gap-junction coupled bursting pancreatic $β $-cells, Chaos, Solitons & Fractals, 68 (2014): 65-71. |
| [46] | [ J. Wang, S. Liu and X. Liu, Bifurcation and firing patterns of the pancreatic $β $-cell ,Int. J Bifurcation Chaos, 9 (2015), 1530024, 11pp. |


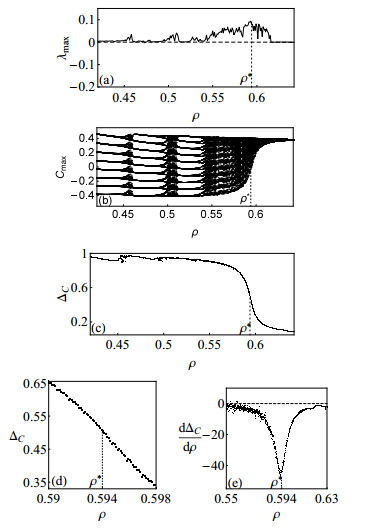
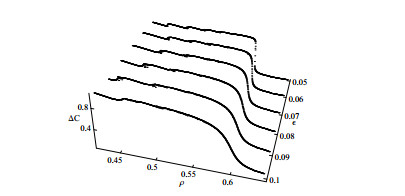
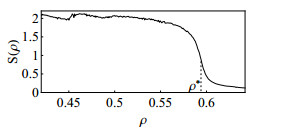
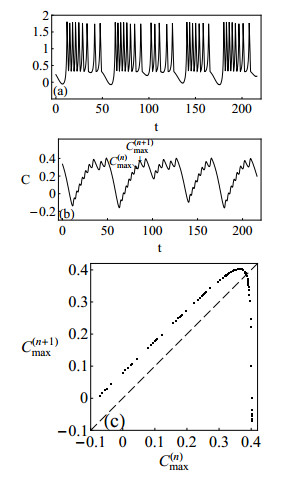

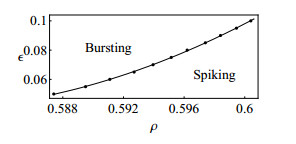
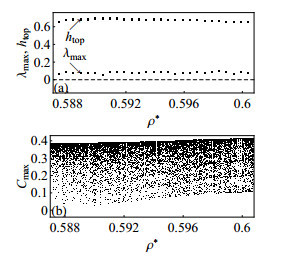
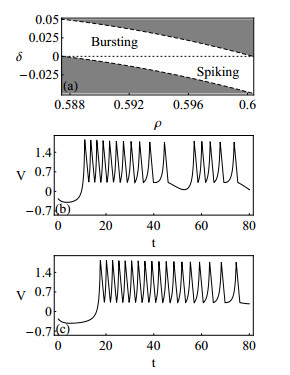
Figures(10)

Jorge Duarte, Cristina Januário, Nuno Martins. A chaotic bursting-spiking transition in a pancreatic beta-cells system: observation of an interior glucose-induced crisis[J]. Mathematical Biosciences and Engineering, 2017, 14(4): 821-842. doi: 10.3934/mbe.2017045







 DownLoad:
DownLoad: