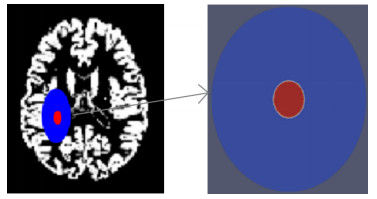
In this study, we introduced a mathematical model mimicking as much as possible the evolutions and interactions between glioma and lactate in the brain, in order to test different therapies and administration protocols. We simulated both glioma cell density evolution and lactate concentration, and considered two therapies: chemotherapy and a treatment targeting lactate production. Three different protocols for administrating the therapies were tested. We compared the efficiency of the combined therapies, depending on the administration protocols and the dosage of the drugs, in order to evaluate the importance of controlling lactate production. Results show that the use of an agent to reduce lactate concentration permits one to significantly reduce the dose of the chemotherapeutic drug.
Citation: Hussein Raad, Cyrille Allery, Laurence Cherfils, Carole Guillevin, Alain Miranville, Thomas Sookiew, Luc Pellerin, Rémy Guillevin. Simulation of tumor density evolution upon chemotherapy alone or combined with a treatment to reduce lactate levels[J]. AIMS Mathematics, 2024, 9(3): 5250-5268. doi: 10.3934/math.2024254
In this study, we introduced a mathematical model mimicking as much as possible the evolutions and interactions between glioma and lactate in the brain, in order to test different therapies and administration protocols. We simulated both glioma cell density evolution and lactate concentration, and considered two therapies: chemotherapy and a treatment targeting lactate production. Three different protocols for administrating the therapies were tested. We compared the efficiency of the combined therapies, depending on the administration protocols and the dosage of the drugs, in order to evaluate the importance of controlling lactate production. Results show that the use of an agent to reduce lactate concentration permits one to significantly reduce the dose of the chemotherapeutic drug.

| [1] |
J. C. L. Alfonso, K. Talkenberger, M. Seifert, B. Klink, A. Hawkins-Daarud, K. R. Swanson, et al., The biology and mathematical modelling of glioma invasion: a review, J. R. Soc. Interface, 14 (2017), 20170490. https://doi.org/10.1098/rsif.2017.0490 doi: 10.1098/rsif.2017.0490

|
| [2] |
H. Alsayed, H. Fakih, A. Miranville, A. Wehbe, On an optimal control problem describing lactate production inhibition, Appl. Anal., 102 (2023), 1711–1731. https://doi.org/10.1080/00036811.2021.1999418 doi: 10.1080/00036811.2021.1999418
|
| [3] |
A. Aubert, R. Costalat, P. J. Magistretti, L. Pellerin, Brain lactate kinetics: modeling evidence for neuronal lactate uptake upon activation, Proceedings of the National Academy of Sciences of the United States of America, 102 (2005), 16448–16453. https://doi.org/10.1073/pnas.0505427102 doi: 10.1073/pnas.0505427102
|
| [4] |
D. Benjamin, M. N. Hall, Combining metformin with lactate transport inhibitors as a treatment modality for cancer - recommendation proposal, Front. Oncol., 12 (2022), 1034397. https://doi.org/10.3389/fonc.2022.1034397 doi: 10.3389/fonc.2022.1034397
|
| [5] |
M. U. Bogdańska, M. Bodnar, J. Belmonte-Beita, M. Murek, P. Schucht, J. Beck, et al., A mathematical model of low grade gliomas treated with temozolomide and its therapeutical implications, Math. Biosci., 288 (2017), 1–13. https://doi.org/10.1016/j.mbs.2017.02.003 doi: 10.1016/j.mbs.2017.02.003
|
| [6] | F. Bonnans, J.-C. Gilbert, C. Lemaréchal, C. Sagastizábal, Optimisation Numérique: Aspects théoriques et pratiques (Mathématiques et Applications), Springer, 1997. |
| [7] |
H. E. Bridgewater, E. M. Bolitho, I. Romero-Canelón, P. J. Sadler, J. P. C. Coverdale, Targeting lactate metabolism with synergistic combinations of synthetic catalysts and monocarboxylate transporter inhibitors, J. Biol. Inorg. Chem., 28 (2023), 345–353. https://doi.org/10.1007/s00775-023-01994-3 doi: 10.1007/s00775-023-01994-3
|
| [8] |
L. E. B. Cabrales, J. I. Montijano, M. Schonbek, A. R. S. Casta$\tilde{\hbox{n}}$eda, A viscous modified Gompertz model for the analysis of the kinetics of tumors under electrochemical therapy, Mathematics and Computers in Simulations, 151 (2018), 96–110. https://doi.org/10.1016/j.matcom.2018.03.005 doi: 10.1016/j.matcom.2018.03.005
|
| [9] |
A. R. S. Casta$\tilde{\hbox{n}}$eda, J. M. Pozo, E. E. Ramirez-Torres, E. J. R. Oria, S. B. Vaillant, J. I. Montijano, et al., Spatio temporal dynamics of direct current in treated anisotropic tumors, Mathematics and Computers in Simulations, 203 (2023), 609–632. https://doi.org/10.1016/j.matcom.2022.07.004 doi: 10.1016/j.matcom.2022.07.004
|
| [10] |
M. M. Chaumeil, M. Radoul, C. Najac, P. Eriksson, P. Viswanath, M. D. Blough, et al., Hyperpolarized $^13$C MR imaging detects no lactate production in mutant IDH1 gliomas: Implications for diagnosis and response monitoring, Neuroimage Clin., 12 (2016), 180–189. https://doi.org/10.1016/j.nicl.2016.06.018 doi: 10.1016/j.nicl.2016.06.018
|
| [11] |
L. Cherfils, S. Gatti, C. Guillevin, A. Miranville, R. Guillevin, On a tumor growth model with brain lactate kinetics, Math. Med. Biol., 39 (2022), 382–409. https://doi.org/10.1093/imammb/dqac010 doi: 10.1093/imammb/dqac010
|
| [12] |
P. Colli, H. Gomez, G. Lorenzo, G. Marinoschi, A. Reali, E. Rocca, Mathematical analysis and simulation study of a phase-field model of prostate cancer growth with chemotherapy and antiangiogenic therapy effects, Math. Mod. Meth. Appl. Sci., 30 (2020), 1253–1295. https://doi.org/10.1142/S0218202520500220 doi: 10.1142/S0218202520500220
|
| [13] |
H. Gomez, Quantitative analysis of the proliferative-to-invasive transition of hypoxic glioma cells, Integr. Biol., 9 (2017), 257–262. https://doi.org/10.1039/C6IB00208K doi: 10.1039/C6IB00208K
|
| [14] |
C. Guillevin, R. Guillevin, A. Miranville, A. Perillat-Mercerot, Analysis of a mathematical model for brain lactate kinetics, Math. Biosci. Eng., 15 (2018), 1225–1242. https://doi.org/10.3934/mbe.2018056 doi: 10.3934/mbe.2018056
|
| [15] |
S. Halford, G. J. Veal, S. R. Wedge, G. S. Payne, C. M. Bacon, P. Sloan, et al., A Phase I Dose-escalation Study of AZD3965, an Oral Monocarboxylate Transporter 1 Inhibitor, in Patients with Advanced Cancer, Clin. Cancer Res., 29 (2023), 1429–1439. https://doi.org/10.1158/1078-0432.CCR-22-2263 doi: 10.1158/1078-0432.CCR-22-2263
|
| [16] |
P. Jacquet, A. Stéphanou, Metabolic Reprogramming, Questioning, and Implications for Cancer, Biology, 10 (2021), 129. https://doi.org/10.3390/biology10020129 doi: 10.3390/biology10020129
|
| [17] |
S. Jbabdi, E. Mandonnet, H. Duffau, L. Capelle, K. R. Swanson, M. Pélégrini-Issac, et al., Simulation of anisotropic growth of low-grade gliomas using diffusion tensor imaging, Magn. Reson. Med., 54 (2005), 616–624. https://doi.org/10.1002/mrm.20625 doi: 10.1002/mrm.20625
|
| [18] |
S. Kumstel, T. Schreiber, L. Goldstein, J. Stenzel, T. Lindner, M. Joksch, et al., Targeting pancreatic cancer with combinatorial treatment of CPI-613 and inhibitors of lactate metabolism, PLoS ONE, 17 (2022), e0266601. https://doi.org/10.1371/journal.pone.0266601 doi: 10.1371/journal.pone.0266601
|
| [19] |
L. Li, A. Miranville, R. Guillevin, Cahn-Hilliard models for glial cells, Appl. Math. Optim., 84 (2021), 1821–1842. https://doi.org/10.1007/s00245-020-09696-x doi: 10.1007/s00245-020-09696-x
|
| [20] |
L. Longhitano, N. Vicario, S. Forte, C. Giallongo, G. Broggi, R. Caltabiano, et al. Lactate modulates microglia polarization via IGFBP6 expression and remodels tumor microenvironment in glioblastoma, Cancer Immunol. Immun., 72 (2023), 1–20. https://doi.org/10.1007/s00262-022-03215-3 doi: 10.1007/s00262-022-03215-3
|
| [21] |
N. Minami, K. Tanaka, T. Sasayama, E. Kohmura, H. Saya, O. Sampetrean, Lactate Reprograms Energy and Lipid Metabolism in Glucose-Deprived Oxidative Glioma Stem Cells, Metabolites, 11 (2021), 325. https://doi.org/10.3390/metabo11050325 doi: 10.3390/metabo11050325
|
| [22] |
R. A. Noble, H. Thomas, Y. Zhao, L. Herendi, R. Howarth, I. Dragoni, et al., Simultaneous targeting of glycolysis and oxidative phosphorylation as a therapeutic strategy to treat diffuse large B-cell lymphoma, Brit. J. Cancer, 127 (2022), 937–947. https://doi.org/10.1038/s41416-022-01848-w doi: 10.1038/s41416-022-01848-w
|
| [23] | H. Raad, C. Allery, L. Cherfils, R. Guillevin, Optimal control of a model for brain lactate kinetics, Asymptotic Anal., Preprint (2023), 1–32. https://doi.org/10.3233/ASY-221823 |
| [24] | H. Raad, C. Allery, L. Cherfils, A. Miranville, R. Guillevin, Optimal control of therapies on a tumor growth model, submitted for publication. |
| [25] |
P. Sonveaux, F. Végran, T. Schroeder, M. C. Wergin, J. Verrax, Targeting lactate-fueled respiration selectively kills hypoxix tumor cells in mice, J. Clin. Invest., 118 (2008), 3930–3942. https://doi.org/10.1172/JCI36843 doi: 10.1172/JCI36843
|
| [26] |
K. R. Swanson, E. C. Alvord, J. D. Murray, Quantifying efficacy of chemotherapy of brain tumors with homogeneous and heterogeneous drug delivery, Acta Biotheor., 50 (2002), 223–237. https://doi.org/10.1023/A:1022644031905 doi: 10.1023/A:1022644031905
|
| [27] |
K. R. Swanson, R. C. Rostornily, E. C. Alvord, A mathematical modelling tool for predicting survival of individual patients following resection of glioblastoma: a proof of principle, Br. J. Cancer, 98 (2007), 113–119. https://doi.org/10.1038/sj.bjc.6604125 doi: 10.1038/sj.bjc.6604125
|
| [28] |
R. Stupp, W. P. Mason, M. J. van den Bent, M. Weller, B. Fisher, M. J. B. Taphoorn, et al., Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma, The New England journal of Medecine, 352 (2005), 987–996. https://doi.org/10.1056/NEJMoa043330 doi: 10.1056/NEJMoa043330
|


















Figures(19) / Tables(1)

Hussein Raad, Cyrille Allery, Laurence Cherfils, Carole Guillevin, Alain Miranville, Thomas Sookiew, Luc Pellerin, Rémy Guillevin. Simulation of tumor density evolution upon chemotherapy alone or combined with a treatment to reduce lactate levels[J]. AIMS Mathematics, 2024, 9(3): 5250-5268. doi: 10.3934/math.2024254







 DownLoad:
DownLoad: