We derive conditions for a nonholonomic system subject to nonlinear constraints (obeying Chetaev's rule) to preserve a smooth volume form. When applied to affine constraints, these conditions dictate that a basic invariant density exists if and only if a certain 1-form is exact and a certain function vanishes (this function automatically vanishes for linear constraints). Moreover, this result can be extended to geodesic flows for arbitrary metric connections and the sufficient condition manifests as integrability of the torsion. As a consequence, volume-preservation of a nonholonomic system is closely related to the torsion of the nonholonomic connection. Examples of nonlinear/affine/linear constraints are considered.
Citation: William Clark, Anthony Bloch. Existence of invariant volumes in nonholonomic systems subject to nonlinear constraints[J]. Journal of Geometric Mechanics, 2023, 15(1): 256-286. doi: 10.3934/jgm.2023011
We derive conditions for a nonholonomic system subject to nonlinear constraints (obeying Chetaev's rule) to preserve a smooth volume form. When applied to affine constraints, these conditions dictate that a basic invariant density exists if and only if a certain 1-form is exact and a certain function vanishes (this function automatically vanishes for linear constraints). Moreover, this result can be extended to geodesic flows for arbitrary metric connections and the sufficient condition manifests as integrability of the torsion. As a consequence, volume-preservation of a nonholonomic system is closely related to the torsion of the nonholonomic connection. Examples of nonlinear/affine/linear constraints are considered.

| [1] | V. V. Kozlov, On the integration theory of the equations in nonholonomic mechanics, Adv.Mech., 8 (1985), 86–107. |
| [2] |
V. V. Kozlov, Invariant measures of smooth dynamical systems, generalized functions and summation methods, Izv. Math, 80 (2016), 342–358. https://doi.org/10.1070/IM8469 doi: 10.1070/IM8469

|
| [3] |
D. V. Zenkov, A. M. Bloch, J. E. Marsden, The Energy-Momentum Method for the Stability of Nonholonomic Systems, Dynam. Stabil. Syst., 13 (1998), 123–165. https://doi.org/10.1080/02681119808806257 doi: 10.1080/02681119808806257
|
| [4] |
A. Ruina, Nonholonomic stability aspects of piecewise holonomic systems, Rep. Math. Phys., 42 (1998), 91–100. https://doi.org/10.1016/S0034-4877(98)80006-2 doi: 10.1016/S0034-4877(98)80006-2
|
| [5] | A. Katok, B. Hasselblatt, Introduction to the Modern Theory of Dynamical Systems, Cambridge University Press. Encyclopedia of Mathematics and its Applications, 1995. https://doi.org/10.1017/CBO9780511809187 |
| [6] | J. Aaronson, An Introduction to Infinite Ergodic Theory, Mathematical Surveys and Monographs, 50 (1997). https://doi.org/10.1090/surv/050 |
| [7] | O.E. Fernandez, The Hamiltonization of nonholonomic systems and its applications, University of Michigan, 2009. |
| [8] | P. Balseiro, L. Garcia-Naranjo, Gauge transformations, twisted Poisson brackets and hamiltonization of nonholonomic systems. Arch. Rational. Mech. Anal., 205 (2012), 267–310. https://doi.org/10.1007/s00205-012-0512-9 |
| [9] |
I. A. Bizyaev, A. V. Borisov, I. S. Mamaev, Hamiltonization of elementary nonholonomic systems, Russ. J. Math. Phys., 22 (2015), 444–453. https://doi.org/10.1134/S1061920815040032 doi: 10.1134/S1061920815040032
|
| [10] | A.V. Bolsonov, A.V. Borisov, I.S. Mamaev, Hamiltonization of nonholonomic systems in the neighborhood of invariant manifolds Regul. Chaotic. Dyn., 15 (2011), 443–464. https://doi.org/10.1134/S1560354711050030 |
| [11] |
A.V. Borisov, I.S. Mamaev, I.A. Bizyaev, The Hierarchy of Dynamics of a Rigid Body Rolling without Slipping and Spinning on a Plane and a Sphere, Regul. Chaotic. Dyn, 18 (2013), 266–328. https://doi.org/10.1134/S1560354713030064 doi: 10.1134/S1560354713030064
|
| [12] |
V.V. Kozlov, On the Integration Theory of Equations of Nonholonomic Mechanics, Regul. Chaotic. Dyn, 7 (2002), 161–176. https://doi.org/10.1070/RD2002v007n02ABEH000203 doi: 10.1070/RD2002v007n02ABEH000203
|
| [13] |
B. Jovanovic, Note on a ball rolling over a sphere: integrable Chaplygin system with an invariant measure without Chaplygin Hamiltonization, Theor. Appl. Mec., 46 (2019), 97–108. https://doi.org/10.2298/TAM190322003J doi: 10.2298/TAM190322003J
|
| [14] |
F. Cantrijn, J. Cortés, M. de León, M. de Diego, On the geometry of generalized Chaplygin systems, Math. Proc. Camb. Phil. Soc., 132 (2002), 323–351. https://doi.org/10.1017/S0305004101005679 doi: 10.1017/S0305004101005679
|
| [15] |
L.C. García-Naranjo, J.C. Marrero, The geometry of nonholonomic Chaplygin systems revisited, Nonlinearity, 33 (2020), 1297–1341. https://doi.org/10.1088/1361-6544/ab5c0a doi: 10.1088/1361-6544/ab5c0a
|
| [16] |
I. Iliyev, On the conditions for the existence of the reducing chaplygin factor, J. Appl. Mathp. Mec., 49 (1985), 295–301. https://doi.org/10.1016/0021-8928(85)90026-7 doi: 10.1016/0021-8928(85)90026-7
|
| [17] |
J. Koiller, Reduction of some classical non-holonomic systems with symmetry, Arch. Ration. Mech. An., 118 (1992), 113–148. https://doi.org/10.1007/BF00375092 doi: 10.1007/BF00375092
|
| [18] | J. C. Monforte, Geometric, control and numerical aspects of nonholonomic systems, Springer-Verlag Berlin Heidelberg, 2004. |
| [19] | J. I. Neimark, N. A. Fufaev, Dynamics of Nonholonomic Systems, American Mathematical Society. Translations of mathematical monographs, 1972. |
| [20] | A.M. Bloch, J. Baillieul, P. Crouch, J.E. Marsden, D. Zenkov, P.S. Krishnaprasad, et al., Nonholonomic Mechanics and Control Springer New York, 2015. https://doi.org/10.1007/978-1-4939-3017-3 |
| [21] |
B. Jovanovic, Non-holonomic geodesic flows on Lie groups and the integrable Suslov problem on SO(4), J. Phys. A-math. Men., 31 (1998), 1415–1422. https://doi.org/10.1088/0305-4470/31/5/011 doi: 10.1088/0305-4470/31/5/011
|
| [22] |
A.M. Bloch, J.E. Marsden, D.V. Zenkov, Quasivelocities and symmetries in non-holonomic systems, Dynam. syst., 24 (2009), 187–222. https://doi.org/10.1080/14689360802609344 doi: 10.1080/14689360802609344
|
| [23] |
D.V. Zenkov, A.M. Bloch, Invariant measures of nonholonomic flows with internal degrees of freedom, Nonlinearity, 16 (2003), 1793–1807. https://doi.org/10.1088/0951-7715/16/5/313 doi: 10.1088/0951-7715/16/5/313
|
| [24] |
Y. N. Federov, L. C. García-Naranjo, J. C. Marrero, Unimodularity and Preservation of Volumes in Nonholonomic Mechanics, J. Nonlinear. Sci., 25 (2015), 203–246. https://doi.org/10.1007/s00332-014-9227-4 doi: 10.1007/s00332-014-9227-4
|
| [25] |
A. Yoshida, P. Morrison, Deformation of Lie-Poisson algebra and chirality, J. Math. Phys., 61 (2020), 092901. https://doi.org/10.1063/1.5145218 doi: 10.1063/1.5145218
|
| [26] |
A. V. Bolsinov, I. S. Mamaev, The Nonexistence of an Invariant Measure for an Inhomogeneous Ellipsoid Rolling on a Plane, Math. Notes, 77 (2005), 855–857. https://doi.org/10.1007/s11006-005-0085-0 doi: 10.1007/s11006-005-0085-0
|
| [27] |
L. C. García-Naranjo, J. C. Marrero, Non-existence of an invariant measure for a homogeneous ellipsoid rolling on the plane, Regul. Chaotic. Dyn, 25 (2013), 372–379. https://doi.org/10.1134/S1560354713040047 doi: 10.1134/S1560354713040047
|
| [28] |
C. J. Blackall, On volume integral invariants of non-holonomic dynamical systems, Am. J. Math., 63 (1941), 155–168. https://doi.org/10.2307/2371286 doi: 10.2307/2371286
|
| [29] | W. Clark, Invariant Measures, Geometry, and Control of Hybrid and Nonholonomic Dynamical Systems, University of Michigan, 2020. |
| [30] |
C. M. Marle, Various approaches to conservative and nonconservative nonholonomic systems, Rep. Math. Phys., 42 (1998), 211–229. https://doi.org/10.1016/S0034-4877(98)80011-6 doi: 10.1016/S0034-4877(98)80011-6
|
| [31] |
M. de León, J.C. Marrero, D.M. de Diego, Mechanical systems with nonlinear constraints, Int. J. Theor. Phys., 36 (1997), 979–995. https://doi.org/10.1007/BF02435796 doi: 10.1007/BF02435796
|
| [32] | A. J. Van Der Schaft, B. M. Maschke, On the Hamiltonian formulation of nonholonomic mechanical systems Rep. Math. Phys., 34 (1994), 225–233. https://doi.org/10.1016/0034-4877(94)90038-8 |
| [33] | A. Isidori, Nonlinear Control Systems, Springer-Verlag London. Communications and Control Engineering, 1995. https://doi.org/10.1007/978-1-84628-615-5 |
| [34] | F. Fassò, L. C. García-Narango, N. Sansonetto, Moving energies as first integrals of nonholonomic systems with affine constraints, Nonlinearity 31 (2018), 755–782. https://doi.org/10.1088/1361-6544/aa9837 |
| [35] | R. Abraham, J. E. Marsden, Foundations of Mechanics, AMS Chelsea publishing. AMS Chelsea Pub./American Mathematical Society, 2008. https://doi.org/10.1090/chel/364 |
| [36] |
G. Forni, The cohomological equation for area-preserving flows on compact surfaces, Electron. Res. Announc., 1 (1995), 114–123. https://doi.org/10.1090/S1079-6762-95-03005-8 doi: 10.1090/S1079-6762-95-03005-8
|
| [37] |
A.N. Livšic, Cohomology of dynamical systems, Mathematica of the USSR-Izvestiya, 6 (1972), 1278–1301. https://doi.org/10.1070/IM1972v006n06ABEH001919 doi: 10.1070/IM1972v006n06ABEH001919
|
| [38] |
L.C. García-Naranjo, A.J. Maciejewski, J.C. Marrero, M. Przybylska, The inhomogeneous Suslov problem, Phy. Lett. A., 378 (2014), 2389–2395. https://doi.org/10.1016/j.physleta.2014.06.026 doi: 10.1016/j.physleta.2014.06.026
|
| [39] | A. M. Vershik, L. D. Faddeev, Lagrangian Mechanics in Invariant Form, Selecta. Math. Sov., 4 (1981), 339–350. |
| [40] |
I. A. Bizyaev, A. V. Borisov, I. S. Mamaev, An Invariant Measure and the Probability of a Fall in the Problem of an Inhomogeneous Disk Rolling on a Plane, Regul. Chaotic. Dyn., 23 (2018), 665–684. https://doi.org/10.1134/S1560354718060035 doi: 10.1134/S1560354718060035
|
| [41] | S. A. Chaplygin, On a rolling of a sphere on a horizontal plane, Mathematical Collection of the Moscow Mathematical Society, 24 (1903), 139–168. |
| [42] | I. A. Bizyaev, A. V. Borisov, I. S. Mamaev, Dynamics of the Chaplygin ball on a rotating plane. Russ. J. Math. Phys., 25 (2018), 423–433. https://doi.org/10.1134/S1061920818040027 |
| [43] |
D. Schneider, Nonholonomic Euler-Poincaré equations and stability in Chaplygin's sphere, Dynam. Syst., 17 (2002), 87–130. https://doi.org/10.1080/02681110110112852 doi: 10.1080/02681110110112852
|
| [44] | A. G. Rojo, A. M. Bloch, Nonholonomic double-bracket equations and the Gauss thermostat, Phys. Rev. E., 80 (2009). https://doi.org/10.1103/PhysRevE.80.025601 |
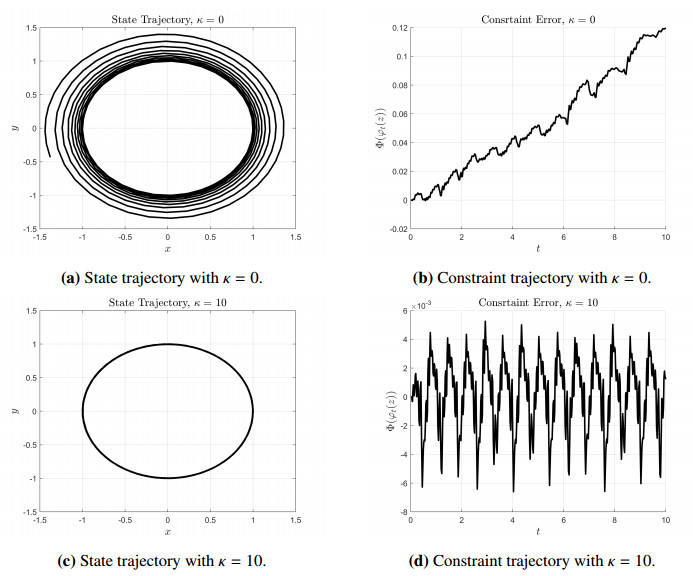

Figures(2)
William Clark, Anthony Bloch. Existence of invariant volumes in nonholonomic systems subject to nonlinear constraints[J]. Journal of Geometric Mechanics, 2023, 15(1): 256-286. doi: 10.3934/jgm.2023011







 DownLoad:
DownLoad: