A double-time-delay Holling Ⅱ predator model with weak Allee effect and age structure was studied in this paper. First, the model was converted into an abstract Cauchy problem. We also discussed the well-posedness of the model and the existence of the equilibrium solution. We analyzed the global stability of boundary equilibrium points, the local stability of positive equilibrium points, and the conditions of the Hopf bifurcation for the system. The conclusion was verified by numerical simulation.
Citation: Yanhe Qiao, Hui Cao, Guoming Xu. A double time-delay Holling Ⅱ predation model with weak Allee effect and age-structure[J]. Electronic Research Archive, 2024, 32(3): 1749-1769. doi: 10.3934/era.2024080
A double-time-delay Holling Ⅱ predator model with weak Allee effect and age structure was studied in this paper. First, the model was converted into an abstract Cauchy problem. We also discussed the well-posedness of the model and the existence of the equilibrium solution. We analyzed the global stability of boundary equilibrium points, the local stability of positive equilibrium points, and the conditions of the Hopf bifurcation for the system. The conclusion was verified by numerical simulation.

| [1] |
M. A. Aziz-Alaoui, M. D. Okiye, Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type Ⅱ schemes, Appl. Math. Lett., 16 (2003), 1069–1075. https://doi.org/10.1016/S0893-9659(03)90096-6 doi: 10.1016/S0893-9659(03)90096-6

|
| [2] |
F. A. Rihan, H. J. Alsakaji, Stochastic delay differential equations of three-species prey-predator system with cooperation among prey species, Discrete Contin. Dyn. Syst. - Ser. S, 15 (2022), 245–263. https://doi.org/10.3934/dcdss.2020468 doi: 10.3934/dcdss.2020468
|
| [3] |
Y. H. Fan, W. T. Li, Permanence for a delayed discrete ratio-dependent predator–prey system with Holling type functional response, J. Math. Anal. Appl., 299 (2004), 357–374. https://doi.org/10.1016/j.jmaa.2004.02.061 doi: 10.1016/j.jmaa.2004.02.061
|
| [4] |
Y. Zhang, Q. L. Zhang, Dynamical analysis of a delayed singular prey–predator economic model with stochastic fluctuations, Complexity, 19 (2014), 23–29. https://doi.org/10.1002/cplx.21486 doi: 10.1002/cplx.21486
|
| [5] |
U. Das, T. K. Kar, Bifurcation analysis of a delayed predator-prey model with Holling type Ⅲ functional response and predator harvesting, J. Nonlinear Dyn., 2014 (2014), 543041. https://doi.org/10.1155/2014/543041 doi: 10.1155/2014/543041
|
| [6] |
Q. B. Gao, N. Olgac, Bounds of imaginary spectra of LTI systems in the domain of two of the multiple time delays, Automatica, 72 (2016), 235–241. https://doi.org/10.1016/j.automatica.2016.05.011 doi: 10.1016/j.automatica.2016.05.011
|
| [7] |
J. Z. Cai, Q. B. Gao, Y. F. Liu, A. G. Wu, Generalized dixon resultant for strong delay-independent stability of linear systems with multiple delays, IEEE Trans. Autom. Control, 1 (2023), 1–8. https://doi.org/10.1109/TAC.2023.3337691 doi: 10.1109/TAC.2023.3337691
|
| [8] |
P. Yang, Hopf bifurcation of an age-structured prey Cpredator model with Holling type Ⅱ functional response incorporating a prey refuge, Nonlinear Anal. Real World Appl., 49 (2019), 368–385. https://doi.org/10.1016/j.nonrwa.2019.03.014 doi: 10.1016/j.nonrwa.2019.03.014
|
| [9] |
D. X. Yan, Y. Cao, Y. Yuan, Stability and Hopf bifurcation analysis of a delayed predator–prey model with age-structure and Holling Ⅲ functional response, Z. Angew. Math. Phys., 74 (2023), 148–172. https://doi.org/10.1007/s00033-023-02036-3 doi: 10.1007/s00033-023-02036-3
|
| [10] |
D. X. Yan, Y. Yuan, X. L. Fu, Asymptotic analysis of an age-structured predator-prey model with ratio-dependent Holling Ⅲ functional response and delays, Evol. Equations Control Theory, 12 (2023), 391–414. https://doi.org/10.3934/eect.2022034 doi: 10.3934/eect.2022034
|
| [11] |
G. Zhu, J. J. Wei, Global stability and bifurcation analysis of a delayed predator-prey system with prey immigration, Electron. J. Qual. Theory Differ. Equations, 13 (2016), 1–20. https://doi.org/10.14232/ejqtde.2016.1.13 doi: 10.14232/ejqtde.2016.1.13
|
| [12] |
L. J. Wang, C. J. Dai, M. Zhao, Hopf bifurcation in an age-structured prey-predator model with Holling Ⅲ response function, Math. Biosci. Eng., 18 (2021), 3144–3159. https://doi.org/10.3934/mbe.2021156 doi: 10.3934/mbe.2021156
|
| [13] |
X. M. Zhang, Z. H. Liu, Periodic oscillations in age-structured ratio-dependent predator–prey model with Michaelis–Menten type functional response, Physica D, 389 (2019), 51–63. https://doi.org/10.1016/j.physd.2018.10.002 doi: 10.1016/j.physd.2018.10.002
|
| [14] |
N. N. Li, W. X. Sun, S. Q. Liu, A stage-structured predator-prey model with Crowley-Martin functional response, Discrete Contin. Dyn. Syst. - Ser. B, 28 (2023), 2463–2489. https://doi.org/10.3934/dcdsb.2022177 doi: 10.3934/dcdsb.2022177
|
| [15] |
B. T. Mulugeta, L. P. Yu, Q. G. Yuan, J. L. Ren. Bifurcation analysis of a predator-prey model with strong Allee effect and Beddington-DeAngelis functional response, Discrete Contin. Dyn. Syst. - Ser. B, 28 (2023), 1938–1963. https://doi.org/10.3934/dcdsb.2022153 doi: 10.3934/dcdsb.2022153
|
| [16] |
C. A. Ibarra, J. Flores, Dynamics of a Leslie-Gower predator-prey model with Holling type Ⅱ functional response, Allee effect and a generalist predator, Math. Comput. Simul., 188 (2021), 1–22. https://doi.org/10.1016/j.matcom.2021.03.035 doi: 10.1016/j.matcom.2021.03.035
|
| [17] |
H. Y. Wang, S. J. Guo, S. Z. Li, Stationary solutions of advective Lotka–Volterra models with a weak Allee effect and large diffusion, Nonlinear Anal. Real World Appl., 56 (2020), 103171. https://doi.org/10.1016/j.nonrwa.2020.103171 doi: 10.1016/j.nonrwa.2020.103171
|
| [18] |
M. H. Wang, M. Kot, Speeds of invasion in a model with strong or weak Allee effects, Math. Biosci., 171 (2001), 83–97. https://doi.org/10.1016/S0025-5564(01)00048-7 doi: 10.1016/S0025-5564(01)00048-7
|
| [19] | P. Magal, S. G. Ruan, Theory and Applications of Abstract Semilinear Cauchy Problems, Springer Cham, Switzerland, 2018. https://doi.org/10.1007/978-3-030-01506-0 |
| [20] | A. Pazy, Semigroups of Linear Operators and Applications to Partial Differential Equations, Springer-Verlag, New York, 1983. https://doi.org/10.1007/978-1-4612-5561-1 |
| [21] | P. Magal, Compact attractors for time-periodic age-structured population models, Electron. J. Differ. Equations, 65 (2001), 1–35. |
| [22] | P. Magal, S. G. Ruan, Infinite Dimensional Dynamical Systems, Springer, New York, 2013. https://doi.org/10.1007/978-1-4614-4523-4 |
| [23] |
H. R. Thieme, Convergence results and a Poincare-Bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755–763. https://doi.org/10.1007/BF00173267 doi: 10.1007/BF00173267
|
| [24] |
Z. H. Liu, P. Magal, S. G. Ruan, Hopf bifurcation for non-densely defined Cauchy problems, Z. Angew. Math. Phys., 62 (2011), 191–222. https://doi.org/10.1007/s00033-010-0088-x doi: 10.1007/s00033-010-0088-x
|
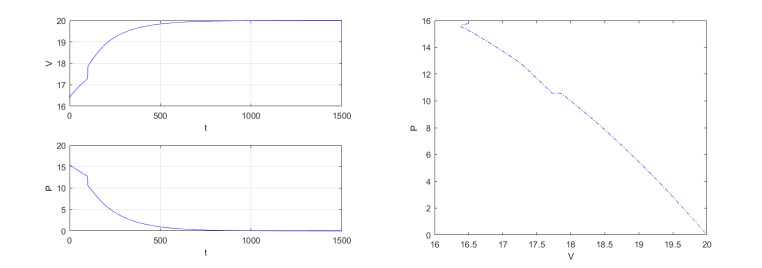
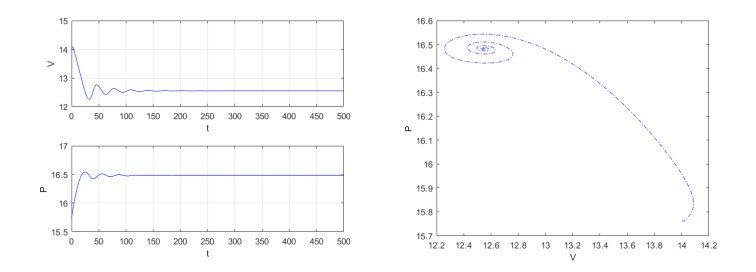
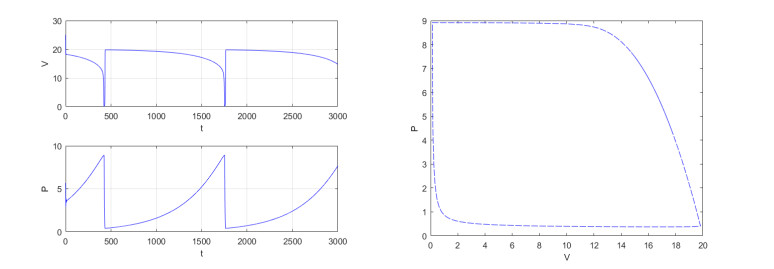
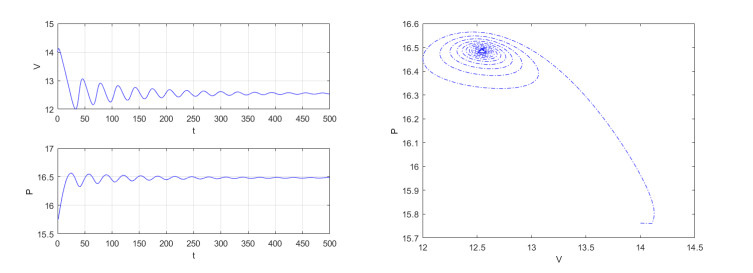
Figures(4)
Yanhe Qiao, Hui Cao, Guoming Xu. A double time-delay Holling Ⅱ predation model with weak Allee effect and age-structure[J]. Electronic Research Archive, 2024, 32(3): 1749-1769. doi: 10.3934/era.2024080







 DownLoad:
DownLoad: