Fly communities exhibit rich ecological dynamics, and one of the important influencing factors is the interaction between species. A discrete Nicholson-type system with multiple time varying delays which considers the mutualism relationship between two fly species is investigated in this paper. Sufficient conditions for the existence of positive periodic solutions are elucidated. The result is obtained by the well-known continuation theorem of coincidence degree theory. An example is attached to illustrate our result. Moreover, the actual biological descriptions obtained from our main result are explained.
Citation: Xinning Niu, Huixin Liu, Dan Li, Yan Yan. Positive periodic solutions for discrete Nicholson system with multiple time-varying delays[J]. Electronic Research Archive, 2023, 31(11): 6982-6999. doi: 10.3934/era.2023354
Fly communities exhibit rich ecological dynamics, and one of the important influencing factors is the interaction between species. A discrete Nicholson-type system with multiple time varying delays which considers the mutualism relationship between two fly species is investigated in this paper. Sufficient conditions for the existence of positive periodic solutions are elucidated. The result is obtained by the well-known continuation theorem of coincidence degree theory. An example is attached to illustrate our result. Moreover, the actual biological descriptions obtained from our main result are explained.

| [1] | G. R. Mullen, L. A. Durden, Medical and Veterinary Entomology, Academic Press, $3^rd$ edition, London, 2019. |
| [2] | D. Calibeo-Hayes, S. S. Denning, S. M. Stringham, J. S. Guy, L. G. Smith, D. W. Watson, Mechanical transmission of Turkey coronavirus by domestic houseflflies (Musca domestica Linnaeaus), Avian Dis., 47 (2003), 149–153. |
| [3] |
F. Francesconi, O. Lupib, Myiasis, Clin. Microbiol. Rev., 25 (2012), 79–105. https://doi.org/10.1128/cmr.00010-11 doi: 10.1128/cmr.00010-11

|
| [4] |
K. Sawabe, K. Tanabayashi, A. Hotta, K. Hoshino, H. Isawa, T. Sasaki, et al., Survival of avian H5N1 inflfluenza a viruses in calliphora nigribarbis (Diptera: Calliphoridae), J. Med. Entomol., 46 (2009), 852–855. https://doi.org/10.1603/033.046.0416 doi: 10.1603/033.046.0416
|
| [5] |
U. B. Pandey, C. D. Nichols, Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery, Pharmacol Rev., 63 (2011), 411–436. https://doi.org/10.1124/pr.110.003293 doi: 10.1124/pr.110.003293
|
| [6] |
A. Jeibmann, W. Paulus, Drosophila melanogaster as a Model Organism of Brain Diseases, Int. J. Mol. Sci., 10 (2009), 407–440. https://doi.org/10.3390/ijms10020407 doi: 10.3390/ijms10020407
|
| [7] |
P. Jagers, The growth and stabilization of populations, Stat. Sci., 6 (1991), 269–274. https://doi.org/10.1214/ss/1177011694 doi: 10.1214/ss/1177011694
|
| [8] |
G. Albano, V. Giorno, P. Román-Román, F. Torres-Ruiz, Study of a general growth model, Commun. Nonlinear Sci. Numer. Simul., 107 (2022), 106100. https://doi.org/10.1016/j.cnsns.2021.106100 doi: 10.1016/j.cnsns.2021.106100
|
| [9] |
L. Berezansky, L. Idels, L. Troib, Global dynamics of Nicholson-type delay systems with applications, Nonlinear Anal. Real World Appl., 12 (2011), 436–445. https://doi.org/10.1016/j.nonrwa.2010.06.028 doi: 10.1016/j.nonrwa.2010.06.028
|
| [10] |
W. Gurney, S. Blythe, R. Nisbet, Nicholson's blowflies revisited, Nature, 287 (1980), 17–21. https://doi.org/10.1038/287017a0 doi: 10.1038/287017a0
|
| [11] |
A. J. Nicholson, An outline of the dynamics of animal populations, Aust. J. Zool., 2 (1954), 9–65. https://doi.org/10.1071/ZO9540009 doi: 10.1071/ZO9540009
|
| [12] |
A. J. Nicholson, The self-adjustment of populations to change, Cold Spring Harb. Symp. Quant. Biol., 22 (1957), 153–173. https://doi.org/10.1101/SQB.1957.022.01.017 doi: 10.1101/SQB.1957.022.01.017
|
| [13] |
C. X. Huang, B. W. Liu, C. F. Qian, J. Cao, Stability on positive pseudo almost periodic solutions of HPDCNNs incorporating $D$ operator, Math. Comput. Simul., 190 (2021), 1150–1163. https://doi.org/10.1016/j.matcom.2021.06.027 doi: 10.1016/j.matcom.2021.06.027
|
| [14] |
W. T. Wang, Mean-square exponential input-to-state stability of stochastic fuzzy delayed Cohen-Grossberg neural networks, J. Exp. Theor. Artif. Intell., 190 (2023), 1150–116. https://doi.org/10.1080/0952813X.2023.2165725 doi: 10.1080/0952813X.2023.2165725
|
| [15] |
P. Amster, A. Déboli, Existence of positive $T$-periodic solutions of a generalized Nicholson's blowflies model with a nonlinear harvesting term, Appl. Math. Lett., 25 (2012), 1203–1207. https://doi.org/10.1016/j.aml.2012.02.040 doi: 10.1016/j.aml.2012.02.040
|
| [16] |
J. W. Li, C. X. Du, Existence of positive periodic solutions for a generalized Nicholson's blowflies model, J. Comput. Appl. Math., 221 (2008), 226–233. https://doi.org/10.1016/j.cam.2007.10.049 doi: 10.1016/j.cam.2007.10.049
|
| [17] |
W. T. Wang, Positive periodic solutions of delayed Nicholson's blowflies models with a nonlinear density-dependent mortality term, Appl. Math. Modell., 36 (2012), 4708–4713. https://doi.org/10.1016/j.apm.2011.12.001 doi: 10.1016/j.apm.2011.12.001
|
| [18] |
W. R. Zhao, C. M. Zhu, H. P. Zhu, On positive periodic solution for the delay Nicholson's blowflies model with a harvesting term, Appl. Math. Modell., 36 (2012), 3335–3340. https://doi.org/10.1016/j.apm.2011.10.011 doi: 10.1016/j.apm.2011.10.011
|
| [19] |
C. X. Huang, B. W. Liu, Traveling wave fronts for a diffusive Nicholson's blowflies equation accompanying mature delay and feedback delay, Appl. Math. Lett., 134 (2022), 108321. https://doi.org/10.1016/j.aml.2022.108321 doi: 10.1016/j.aml.2022.108321
|
| [20] |
C. X. Huang, X. D. Ding, Dynamics of the diffusive Nicholson's blowflies equation with two distinct distributed delays, Appl. Math. Lett., 145 (2023), 108741. https://doi.org/10.1016/j.aml.2023.108741 doi: 10.1016/j.aml.2023.108741
|
| [21] |
J. Sugie, Y. Yan, M. Z. Qu, Effect of decimation on positive periodic solutions of discrete generalized Nicholson's blowflies models with multiple time-varying delay, Commun. Nonlinear Sci. Numer. Simul., 97 (2021), 105731. https://doi.org/10.1016/j.cnsns.2021.105731 doi: 10.1016/j.cnsns.2021.105731
|
| [22] |
S. J. Gao, L. S. Chen, Dynamic complexities in a single-species discrete population model with stage structure and birth pulses, Chaos, Solitons Fractals, 23 (2005), 519–527. https://doi.org/10.1016/j.chaos.2004.05.047 doi: 10.1016/j.chaos.2004.05.047
|
| [23] |
J. S. Yu, B. Zheng, Modeling Wolbachia infection in mosquito population via discrete dynamical models, J. Differ. Equations Appl., 25 (2019), 1549–1567. https://doi.org/10.1080/10236198.2019.1669578 doi: 10.1080/10236198.2019.1669578
|
| [24] |
D. J. Gerla, W. M. Mooij, Alternative stable states and alternative endstates of community assembly through intra- and interspecific positive and negative interactions, Theor. Popul. Biol., 96 (2014), 8–18. https://doi.org/10.1016/j.tpb.2014.07.001 doi: 10.1016/j.tpb.2014.07.001
|
| [25] |
J. P. Twining, W. I. Montgomery, D, G. Tosh, Declining invasive grey squirrel populations may persist in refugia as native predator recovery reverses squirrel species replacement, J. Appl. Ecol., 58 (2021), 248–260. https://doi.org/10.1111/1365-2664.13660 doi: 10.1111/1365-2664.13660
|
| [26] |
Q. Zhou, The positive periodic solution for Nicholson-type delay system with linear harvesting terms, Appl. Math. Modell., 37 (2013), 5581–5590. https://doi.org/10.1016/j.apm.2012.10.056 doi: 10.1016/j.apm.2012.10.056
|
| [27] |
P. Amster, A. Déboli, Necessary and sufficient conditions for the existence of periodic solutions of a nicholson type delay system, Differ. Equations Dyn. Syst., 27 (2019), 439–447. https://doi.org/10.1007/s12591-016-0285-y doi: 10.1007/s12591-016-0285-y
|
| [28] |
G. Ossandóna, D. Sepúlveda, Existence and exponential stability of periodic solutions of Nicholson-type systems with nonlinear density-dependent mortality and linear harvesting, Electron. J. Qual. Theory Differ. Equations, 15 (2023), 1–18. https://doi.org/10.14232/ejqtde.2023.1.15 doi: 10.14232/ejqtde.2023.1.15
|
| [29] | R. E. Gaines, J. L. Mawhin, Coincidence Degree, and Nonlinear Differential Equations, Springer, 1977. |
| [30] | J. L. Mawhin, Topological degree methods in nonlinear boundary value problems, in CBMS Regional Conference Series in Mathematics, 1979. |
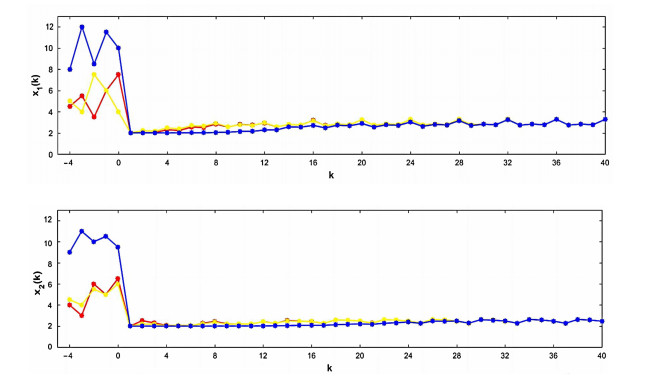
Figures(1)
Xinning Niu, Huixin Liu, Dan Li, Yan Yan. Positive periodic solutions for discrete Nicholson system with multiple time-varying delays[J]. Electronic Research Archive, 2023, 31(11): 6982-6999. doi: 10.3934/era.2023354







 DownLoad:
DownLoad: