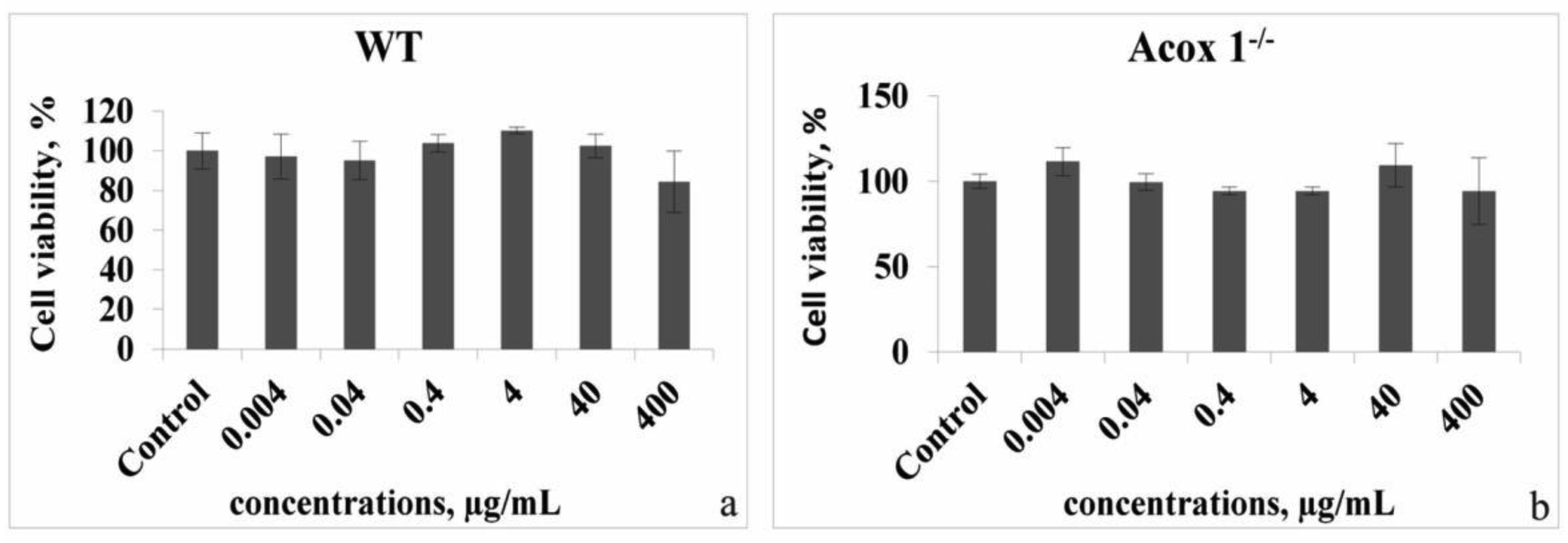
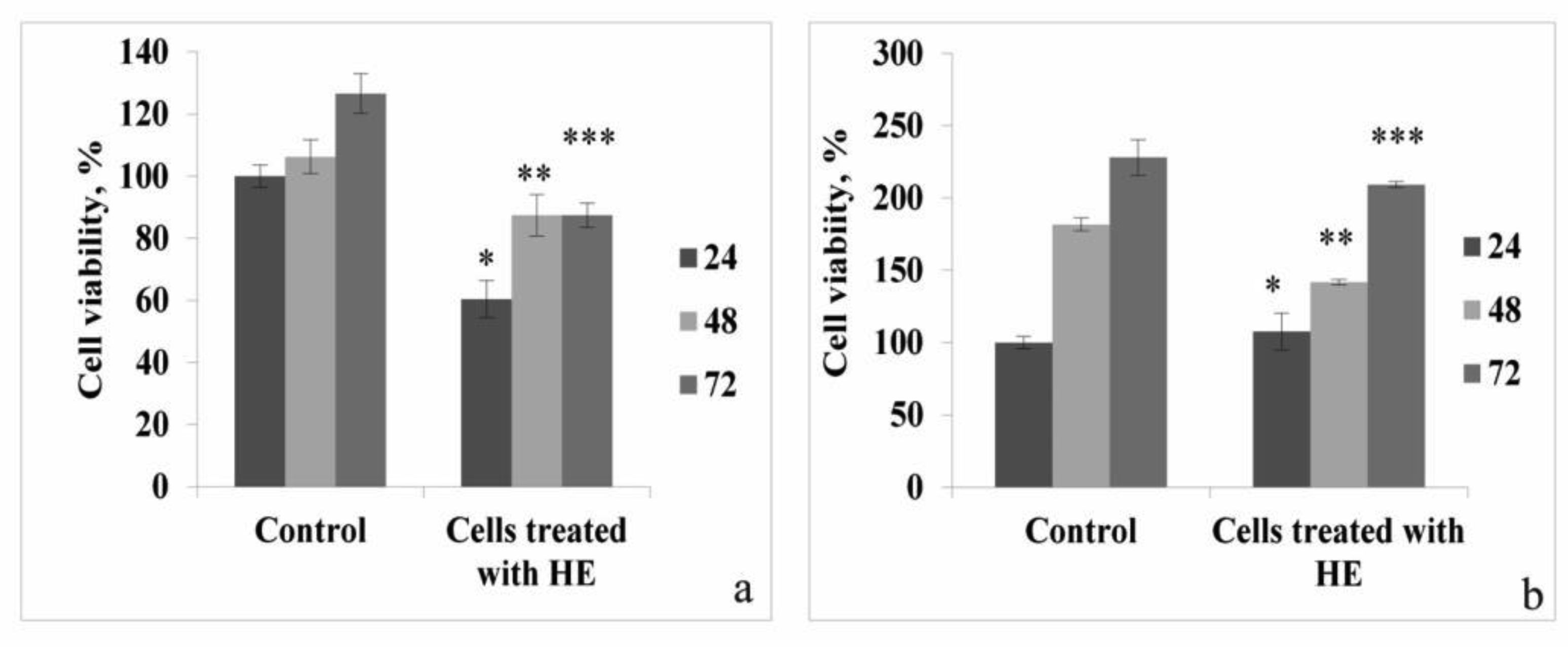
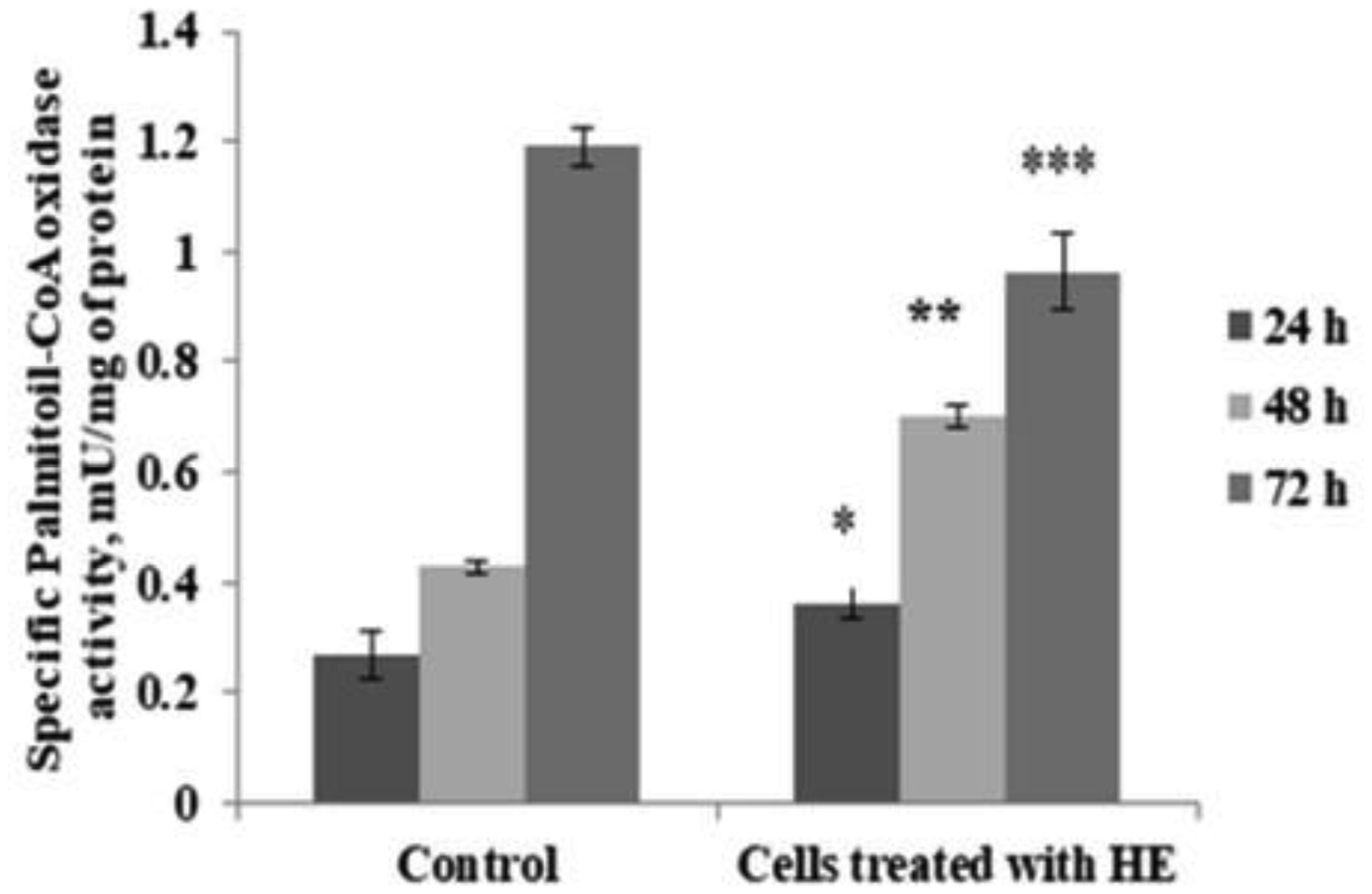
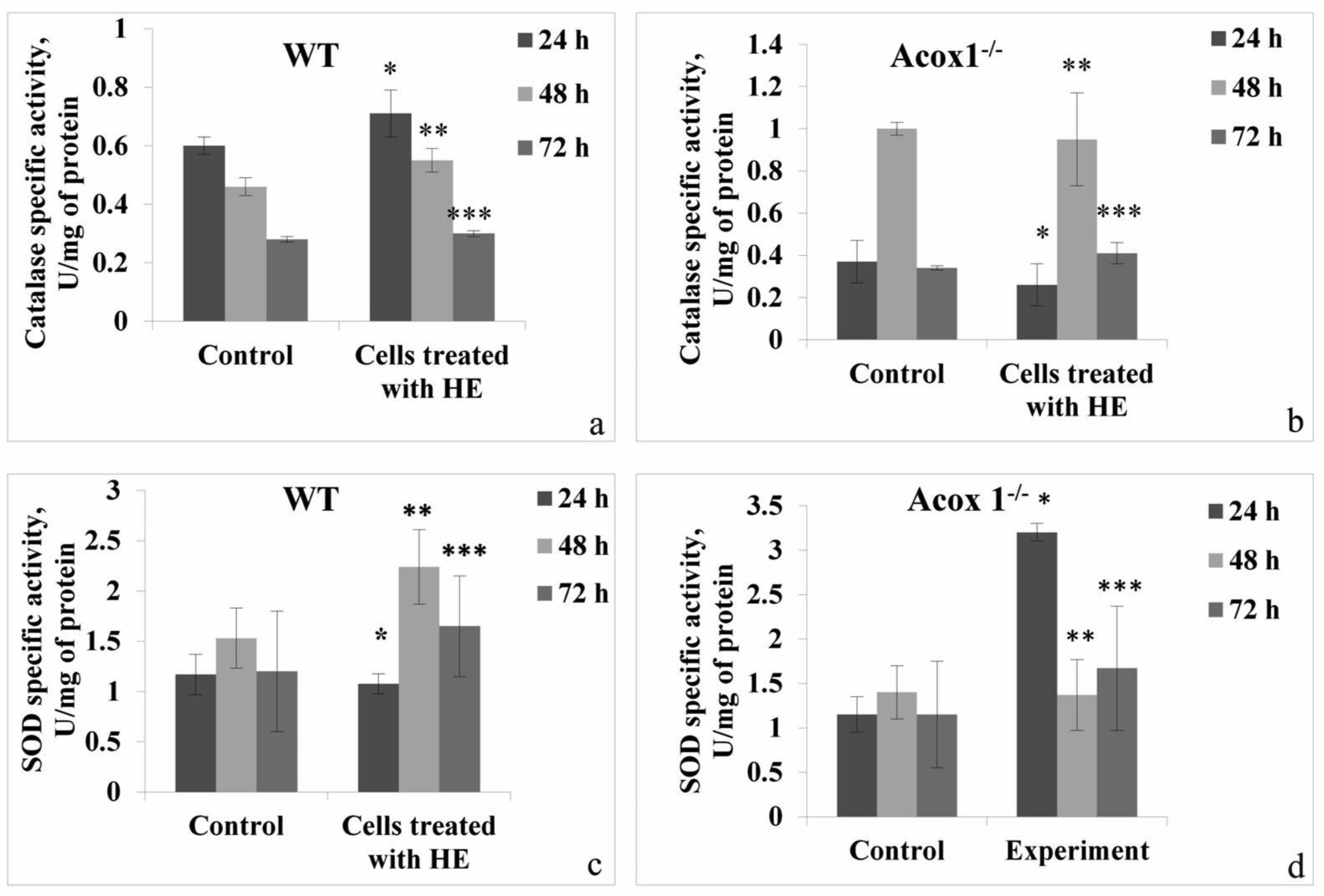
In the presented work, we aimed to investigate the antioxidant and possible neuroprotective capacity of extract of the aerial parts of Hypericum alpestre, found in high altitude Armenian landscape. The neuroprotective activity was evaluated using BV-2 wild type (WT) cells and acyl-CoA oxidase 1 (ACOX1) deficient (Acox1-/-) microglial cell lines. In the chemical-based tests, H. alpestre extract showed high antioxidant activity, which was maintained even after heat treatment at 121 °C for 30 min. MTT test showed that the sub-cytotoxic concentration of investigated extracts for both microglial cell lines was 40 µg/mL. There were no significant changes in catalase activity during all period of treatment in both cell lines, meanwhile, SOD activity increased (up to 30%) in WT cells during the 48 h treatment. Increase of SOD activity (up to 50%) in Acox-/- cells was observed under the 24 h treatment. Significant modulation in activity of palmitoyl-CoA oxidase 1 was noticed only during the 48 h treatment of WT microglial cells. These results evidenced the pro-oxidant activity of the investigated extract. This finding can serve as a basis for further evaluation of plant extracts influence on cancer cell lines.
Citation: Mikayel Ginovyan, Pierre Andreoletti, Mustapha Cherkaoui-Malki, Naira Sahakyan. Hypericum alpestre extract affects the activity of the key antioxidant enzymes in microglial BV-2 cellular models[J]. AIMS Biophysics, 2022, 9(2): 161-171. doi: 10.3934/biophy.2022014
In the presented work, we aimed to investigate the antioxidant and possible neuroprotective capacity of extract of the aerial parts of Hypericum alpestre, found in high altitude Armenian landscape. The neuroprotective activity was evaluated using BV-2 wild type (WT) cells and acyl-CoA oxidase 1 (ACOX1) deficient (Acox1-/-) microglial cell lines. In the chemical-based tests, H. alpestre extract showed high antioxidant activity, which was maintained even after heat treatment at 121 °C for 30 min. MTT test showed that the sub-cytotoxic concentration of investigated extracts for both microglial cell lines was 40 µg/mL. There were no significant changes in catalase activity during all period of treatment in both cell lines, meanwhile, SOD activity increased (up to 30%) in WT cells during the 48 h treatment. Increase of SOD activity (up to 50%) in Acox-/- cells was observed under the 24 h treatment. Significant modulation in activity of palmitoyl-CoA oxidase 1 was noticed only during the 48 h treatment of WT microglial cells. These results evidenced the pro-oxidant activity of the investigated extract. This finding can serve as a basis for further evaluation of plant extracts influence on cancer cell lines.
acyl-CoA oxidase 1
Dulbecco's modified Eagle medium
dimethyl sulfoxide
1,1-diphenyl-2-picrylhydrazyl
ethylenediamine tetraacetic acid
essential oil
fetal bovine serum
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
Phosphate-buffered saline
radioimmunoprecipitation
reactive oxygen species
superoxide dismutase
very-long-chain fatty acids
wild type
gallic acid

| [1] |
Ginovyan M, Petrosyan M, Trchounian A (2017) Antimicrobial activity of some plant materials used in Armenian traditional medicine. BMC Complem Altern M 17: 1-9. https://doi.org/10.1186/s12906-017-1573-y 
|
| [2] |
Ginovyan M, Ayvazyan A, Nikoyan A, et al. (2020) Phytochemical screening and detection of antibacterial components from crude extracts of some Armenian herbs using TLC-bioautographic technique. Curr Microbiol 77: 1223-1232. https://doi.org/10.1007/s00284-020-01929-0
|
| [3] |
Ginovyan M, Trchounian A (2019) Novel approach to combat antibiotic resistance : Evaluation of some Armenian herb crude extracts for their antibiotic modulatory and antiviral properties. J Appl Microbiol 127: 472-480. https://doi.org/10.1111/jam.14335
|
| [4] |
Sahakyan N, Petrosyan M, Koss-Mikołajczyk I, et al. (2019) The Caucasian flora: a still-to-be-discovered rich source of antioxidants. Free Radical Res 53: 1153-1162. https://doi.org/10.1080/10715762.2019.1648799
|
| [5] |
Nürk NM, Uribe-Convers S, Gehrke B, et al. (2015) Oligocene niche shift, Miocene diversification—cold tolerance and accelerated speciation rates in the St. John's Worts (Hypericum, Hypericaceae). BMC Evol Biol 15: 1-13. https://doi.org/10.1186/s12862-015-0359-4
|
| [6] |
Bahmani M, Taherikalani M, Khaksarian M, et al. (2019) Phytochemical profiles and antibacterial activities of hydroalcoholic extracts of Origanum vulgare and Hypericum perforatum and carvacrol and hypericin as a promising anti-Staphylococcus aureus. Mini-Rev Med Chem 19: 923-932. https://doi.org/10.2174/1389557519666190121124317
|
| [7] | Oliveira AI, Pinho C, Sarmento B, et al. (2016) Neuroprotective activity of Hypericum perforatum and its major components. Front Plant Sci 7: 1004. https://doi.org/10.3389/fpls.2016.01004 |
| [8] |
Sytar O, Švedienė J, Ložienė K, et al. (2016) Antifungal properties of hypericin, hypericin tetrasulphonic acid and fagopyrin on pathogenic fungi and spoilage yeasts. Pharm Biol 54: 3121-3125. https://doi.org/10.1080/13880209.2016.1211716
|
| [9] |
Zou Y, Lu Y, Wei D (2004) Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J Agr Food Chem 52: 5032-5039. https://doi.org/10.1021/jf049571r
|
| [10] |
Nahrstedt A, Butterweck V (2010) Lessons learned from herbal medicinal products: the example of St. John's wort. J Nat Prod 73: 1015-1021. https://doi.org/10.1021/np1000329
|
| [11] |
Nahrstedt A, Butterweck V (1997) Biologically active and other chemical constituents of the herb of Hypericum perforation L. Pharmacopsychiatry 30: 129-134. https://doi.org/10.1055/s-2007-979533
|
| [12] | Beerhues L (2011) Biosynthesis of the active Hypericum perforatum constituents. Med Aroma Plant Sci Biotechnol 5: 70-77. |
| [13] |
Linde K (2009) St. John's Wort—an overview. Complement Med Res 16: 146-155. https://doi.org/10.1159/000209290
|
| [14] |
Velingkar VS, Gupta GL, Hegde NB (2017) A current update on phytochemistry, pharmacology and herb—drug interactions of Hypericum perforatum. Phytochem Rev 16: 725-744. https://doi.org/10.1007/s11101-017-9503-7
|
| [15] |
Dauncey EA, Irving JTW, Allkin R (2019) A review of issues of nomenclature and taxonomy of Hypericum perforatum L. and Kew's medicinal plant names services. J Pharm Pharmacol 71: 4-14. https://doi.org/10.1111/jphp.12831
|
| [16] |
Silva BA, Dias ACP, Ferreres F, et al. (2004) Neuroprotective effect of H. perforatum extracts on β-amyloid-induced neurotoxicity. Neurotox Res 6: 119-130. https://doi.org/10.1007/BF03033214
|
| [17] |
Xu WJ, Li RJ, Quasie O, et al. (2016) Polyprenylated tetraoxygenated xanthones from the roots of Hypericum monogynum and their neuroprotective activities. J Nat Prod 79: 1971-1981. https://doi.org/10.1021/acs.jnatprod.6b00251
|
| [18] |
Yan XT, An Z, Huangfu Y, et al. (2019) Polycyclic polyprenylated acylphloroglucinol and phenolic metabolites from the aerial parts of Hypericum elatoides and their neuroprotective and anti-neuroinflammatory activities. Phytochemistry 159: 65-74. https://doi.org/10.1016/j.phytochem.2018.12.011
|
| [19] |
Zeng YR, Li YN, Lou HY, et al. (2021) Polycyclic polyprenylated acylphloroglucinol derivatives with neuroprotective effects from Hypericum monogynum. J Asian Nat Prod Res 23: 73-81. https://doi.org/10.1080/10286020.2019.1698551
|
| [20] | Aparecida M, Cagnin H, Gomes CMR, et al. (1977) Biochemical systematics: methods and principles. Flower Plants. Plant Systematics and Evolution / Entwicklungsgeschichte und Systematik der Pflanzen, Vienna: Springer 53-76. https://doi.org/10.1007/978-3-7091-7076-2_5 |
| [21] |
Ginovyan MM, Sahakyan NZ, Petrosyan MT, et al. (2021) Antioxidant potential of some herbs represented in Armenian flora and characterization of phytochemicals. Proc YSU B Chem Biol Sci 55: 25-38. https://doi.org/10.46991/PYSU:B/2021.55.1.025
|
| [22] | Ginovyan M, Koziara Z, Baranowska M, et al. (2021) Antioxidant properties of some herbal extracts from Armenian flora, evaluation of their cytotoxicity and genotoxicity on HT29 cell line ROS Roundabout—Many Exit Options, Final Meeting of the NutRedOx COST Action CA16112, Gdansk, Poland. 28. https://www.researchgate.net/publication/354792182 |
| [23] |
Sahakyan N, Andreoletti P, Petrosyan M, et al. (2022) Essential oils of basil cultivars selectively affect the activity of antioxidant enzymes in murine glial cells. Curr Nutraceut 3: e171221199126. https://doi.org/10.2174/2665978602666211217143112
|
| [24] |
Singh I, Pujol A (2010) Pathomechanisms underlying X-adrenoleukodystrophy: a three-hit hypothesis. Brain Pathol 20: 838-844. https://doi.org/10.1111/j.1750-3639.2010.00392.x
|
| [25] |
Rojas JJ, Ochoa VJ, Ocampo SA, et al. (2006) Screening for antimicrobial activity of ten medicinal plants used in Colombian folkloric medicine: A possible alternative in the treatment of non-nosocomial infections. BMC Complem Altern M 6: 2. https://doi.org/10.1186/1472-6882-6-2
|
| [26] |
Hambardzumyan S, Sahakyan N, Petrosyan M, et al. (2020) Origanum vulgare L. extract-mediated synthesis of silver nanoparticles, their characterization and antibacterial activities. AMB Express 10: 162. https://doi.org/10.1186/s13568-020-01100-9
|
| [27] | Abdel-Naim AB, Alghamdi AA, Algandaby MM, et al. (2018) Rutin isolated from Chrozophora tinctoria enhances bone cell proliferation and ossification markers. Oxid Med Cell Longev 2018: 5106469. https://doi.org/10.1155/2018/5106469 |
| [28] |
Moghrovyan A, Parseghyan L, Sevoyan G, et al. (2022) Antinociceptive, anti-inflammatory, and cytotoxic properties of Origanum vulgare essential oil, rich with β-caryophyllene and β-caryophyllene oxide. Korean J Pain 35: 140-151. https://doi.org/10.3344/kjp.2022.35.2.140
|
| [29] |
Smith PK, Krohn RI, Hermanson GT, et al. (1985) Measurement of protein using bicinchoninic acid. Anal Biochem 150: 76-85. https://doi.org/10.1016/0003-2697(85)90442-7
|
| [30] |
Malki CM, Bardot O, Lhuguenot JC, et al. (1990) Expression of liver peroxisomal proteins as compared to other organelle marker enzymes in rats treated with hypolipidemic agents. Biol Cell 69: 83-92. https://doi.org/10.1111/j.1768-322X.1990.tb00002.x
|
| [31] |
Oaxaca-Castillo D, Andreoletti P, Vluggens A, et al. (2007) Biochemical characterization of two functional human liver acyl-CoA oxidase isoforms 1a and 1b encoded by a single gene. Biochem Bioph Res Co 360: 314-319. https://doi.org/10.1016/j.bbrc.2007.06.059
|
| [32] |
Beauchamp C, Fridovich I (1971) Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal Biochem 44: 276-287. https://doi.org/10.1016/0003-2697(71)90370-8
|
| [33] |
Judge S, Jang YM, Smith A, et al. (2005) Age-associated increases in oxidative stress and antioxidant enzyme activities in cardiac interfibrillar mitochondria: implications for the mitochondrial theory of aging. FASEB J 19: 1-21. https://doi.org/10.1096/fj.04-2622fje
|
| [34] |
Kraus B, Wolff H, Heilmann J, et al. (2007) Influence of Hypericum perforatum extract and its single compounds on amyloid-beta mediated toxicity in microglial cells. Life Sci 81: 884-894. https://doi.org/10.1016/j.lfs.2007.07.020
|
| [35] |
Sahakyan N, Andreoletti P, Cherkaoui-Malki M, et al. (2021) Artemisia dracunculus L. essential oil phytochemical components trigger the activity of cellular antioxidant enzymes. J Food Biochem 45: e13691. https://doi.org/10.1111/jfbc.13691
|
| [36] |
Danova K, Nikolova-Damianova B, Denev R, et al. (2012) Influence of vitamins on polyphenolic content, morphological development, and stress response in shoot cultures of Hypericum spp. Plant Cell Tiss Org 110: 383-393. https://doi.org/10.1007/s11240-012-0159-0
|
| [37] |
Gioti EM, Fiamegos YC, Skalkos DC, et al. (2009) Antioxidant activity and bioactive components of the aerial parts of Hypericum perforatum L. from Epirus, Greece. Food Chem 117: 398-404. https://doi.org/10.1016/j.foodchem.2009.04.016
|
| [38] |
Attia M, Essa EA, Zaki RM, et al. (2020) An overview of the antioxidant effects of ascorbic acid and alpha lipoic acid (in liposomal forms) as adjuvant in cancer treatment. Antioxidants 9: 359. https://doi.org/10.3390/antiox9050359
|
Figures(4)

Mikayel Ginovyan, Pierre Andreoletti, Mustapha Cherkaoui-Malki, Naira Sahakyan. Hypericum alpestre extract affects the activity of the key antioxidant enzymes in microglial BV-2 cellular models[J]. AIMS Biophysics, 2022, 9(2): 161-171. doi: 10.3934/biophy.2022014







 DownLoad:
DownLoad: