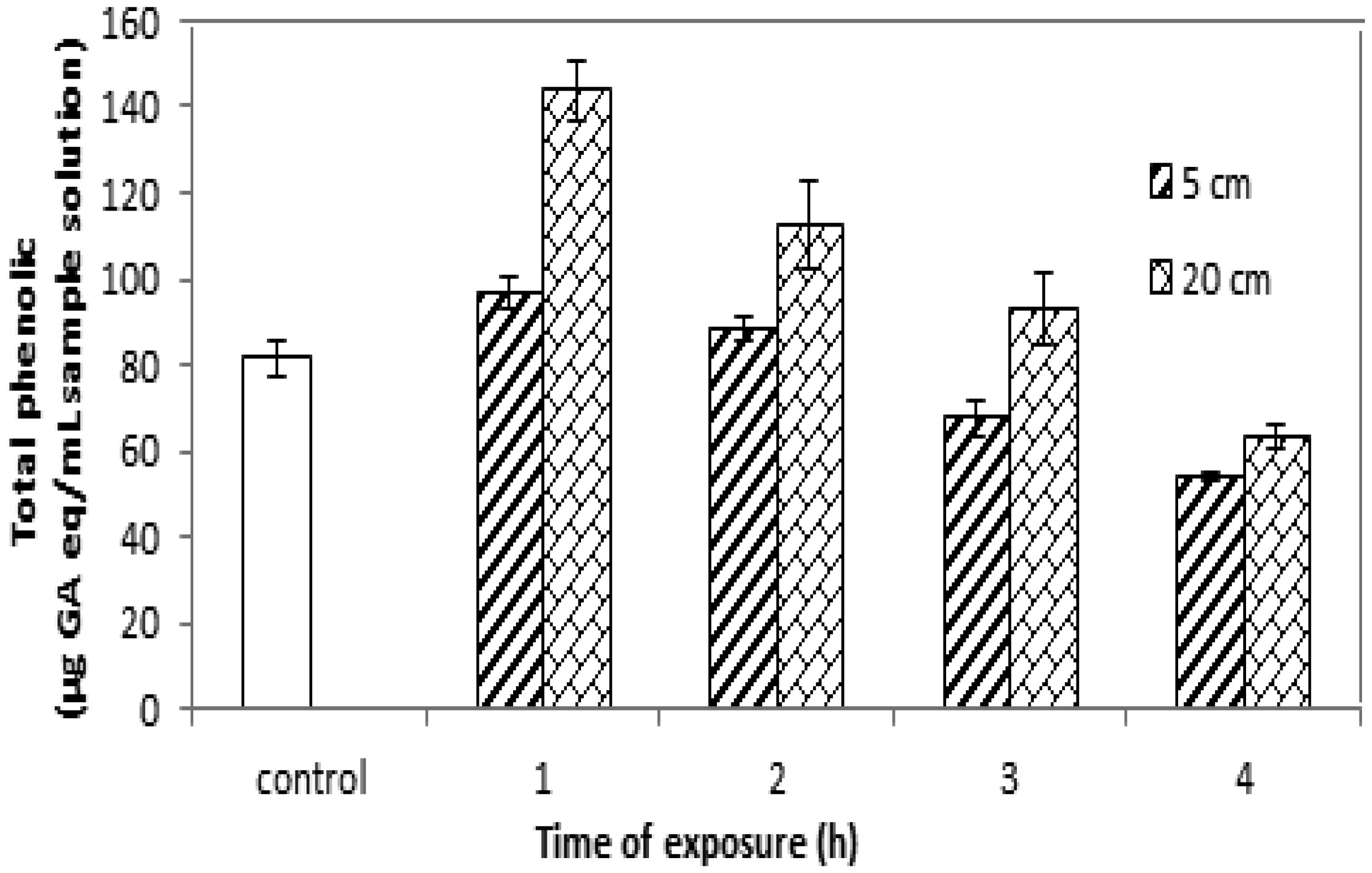
Some seeds are a major source of vitamins and minerals and contain primary and natural antioxidants, making them useful as medical resources to treat infectious diseases in poor and developing countries. This research aimed to study and analyze the effects of ultraviolet A radiation on the molecular structure, chemical composition and photochemical compounds of Salvia hispanica L. seeds (chia seeds). The results show that ultraviolet A radiation significantly affected the arrangement, size, interconnection and orientation of the Salvia hispanica seed molecules. Also, significant changes in the carbohydrate, protein and fat contents were observed, with little variation in the total fibers comprising the Salvia hispanica L. (chia seeds), after exposure to ultraviolet A radiation for different times at dissimilar distances. The phenolic content and flavonoid content in the Salvia hispanica L. seeds varied after exposure to ultraviolet A radiation for 1, 2, 3 and 4 hours at 5-cm and 20-cm distances from the ultraviolet source.
Citation: Reham Ebrahim, Aya Abdelrazek, Hamed El-Shora, Abu Bakr El-Bediwi. Effect of ultraviolet radiation on molecular structure and photochemical compounds of Salvia hispanica medical seeds[J]. AIMS Biophysics, 2022, 9(2): 172-181. doi: 10.3934/biophy.2022015
Some seeds are a major source of vitamins and minerals and contain primary and natural antioxidants, making them useful as medical resources to treat infectious diseases in poor and developing countries. This research aimed to study and analyze the effects of ultraviolet A radiation on the molecular structure, chemical composition and photochemical compounds of Salvia hispanica L. seeds (chia seeds). The results show that ultraviolet A radiation significantly affected the arrangement, size, interconnection and orientation of the Salvia hispanica seed molecules. Also, significant changes in the carbohydrate, protein and fat contents were observed, with little variation in the total fibers comprising the Salvia hispanica L. (chia seeds), after exposure to ultraviolet A radiation for different times at dissimilar distances. The phenolic content and flavonoid content in the Salvia hispanica L. seeds varied after exposure to ultraviolet A radiation for 1, 2, 3 and 4 hours at 5-cm and 20-cm distances from the ultraviolet source.

| [1] |
A.-H.-Mackerness S, Surplus SL, Jordan BR, et al. (1998) Effects of supplementary ultraviolet-B radiation on photosynthetic transcripts at different stages of leaf development and light levels in pea (Pisum sativum L.): Role of active oxygen species and antioxidant enzymes. Photochem Photobiol 68: 88-96. https://doi.org/10.1111/j.1751-1097.1998.tb03257.x 
|
| [2] |
He YY, Häder DP (2002) Involvement of reactive oxygen species in the UV-B damage to the cyanobacterium Anabaena sp. J Photoch Photobio B 66: 73-80. https://doi.org/10.1016/S1011-1344(01)00278-0
|
| [3] | Jain K, Kataria S, Guruprasad KN (2004) Effect of UV-B radiation on antioxidant enzymes and its modulation by benzoquinone and α-tocopherol in cucumber cotyledons. Curr Sci India 87: 87-90. https://www.jstor.org/stable/24107986 |
| [4] |
Zu YG, Pang HH, Yu JH, et al. (2010) Responses in the morphology, physiology and biochemistry of Taxus chinensis var. mairei grown under supplementary UV-B radiation. J Photoch Photobio B 98: 152-158. https://doi.org/10.1016/j.jphotobiol.2009.12.001
|
| [5] |
Tegelberg R, Julkunen-Tiitto R, Aphalo PJ (2001) The effects of long-term elevated UV-B on the growth and phenolics of field-grown silver birch (Betula pendula). Global Change Biol 7: 839-848. https://doi.org/10.1046/j.1354-1013.2001.00453.x
|
| [6] |
Shohael AM, Ali MB, Yu KW, et al. (2006) Effect of light on oxidative stress, secondary metabolites and induction of antioxidant enzymes in eleutherococcus senticosus somatic embryos in bioreactor. Process Biochem 41: 1179-1185. https://doi.org/10.1016/j.procbio.2005.12.015
|
| [7] |
Pu GB, Ma DM, Chen JL, et al. (2009) Salicylic acid activates artemisinin biosynthesis in Artemisia annua L. Plant Cell Rep 28: 1127-1135. https://doi.org/10.1007/s00299-009-0713-3
|
| [8] |
Rai R, Meena RP, Smita SS, et al. (2011) UV-B and UV-C pre-treatments induce physiological changes and artemisinin biosynthesis in Artemisia annua L.—an antimalarial plant. J Photoch Photobio B 105: 216-225. https://doi.org/10.1016/j.jphotobiol.2011.09.004
|
| [9] |
Takshak S, Agrawal SB (2015) Defense strategies adopted by the medicinal plant Coleus forskohlii against supplemental ultraviolet-B radiation: augmentation of secondary metabolites and antioxidants. Plant Physiol Bioch 97: 124-138. https://doi.org/10.1016/j.plaphy.2015.09.018
|
| [10] | El-Bediwi AB, Hasanin S, Abdelrazek A, et al. (2018) Influence of UVC on growth behavior, internal structure, enzymes and free radical of Nigella Sativa plant. Res Rev Biosic 13: 142. |
| [11] | El-Bediwi AB, Hasanin S, Abdelrazek A, et al. (2018) Effect of ultraviolet on morphological and secondary metabolites content of garden cress. Int J Sci Res Sci Eng Techn 4: 187-194. |
| [12] | El-Bediwi AB, Hager Y, El-Shora HM (2020) UVC radiation effects on the internal structure and medical contents of ammimajus. Int J Biotech & Bioeng 6: 7-21. |
| [13] | El-Bediwi AB, Hager Y, El-Shora HM (2021) Effect of Ultraviolet radiation on growth, structure, and bio-contents of Foeniculum vulgare. Int J Med Res Health Sci 10: 94-102. |
| [14] | Chen YH, Guo QS, Liu L, et al. (2011) Influence of fertilization and drought stress on the growth and production of secondary metabolites in Prunella vulgaris L. J Med Plants Res 5: 1749-1755. https://doi.org/10.5897/JMPR.9000792 |
| [15] | Wu Z, Guo Q, Wang Q, et al. (2010) Effects of lead, copper and cadmium stresses on growth and inherent quality of Prunalla vulgaris. China J Chin Mater Med 35: 263-267. https://doi.org/10.4268/cjcmm20100301 |
| [16] |
Chen Y, Yu M, Zhu Z, et al. (2013) Optimisation of potassium chloride nutrition for proper growth, physiological development and bioactive component production in Prunella vulgaris L. Plos One 8: e66259. https://doi.org/10.1371/journal.pone.0066259
|
| [17] |
Attard E (2013) A rapid microtitre plate Folin-Ciocalteu method for the assessment of polyphenols. Open Life Sci 8: 48-53. https://doi.org/10.2478/s11535-012-0107-3
|
| [18] | Kiranmai M, Kumar CBM, Mohammed I (2011) Comparison of total flavanoid content of Azadirachta indica root bark extracts prepared by different methods of extraction. Res J Pharm Biol Chem Sci 2: 254-261. http://rjpbcs.com/pdf/2011_2(3)/31.pdf |
| [19] | Horwitz W, Fisher HJ, Robertson AH, et al. (1955) Official methods of analysis of the association of official agricultural chemists. Association of Official Analytical Chemists . Washington, D.C.: . https://doi.org/10.1093/clinchem/2.5.375a |
| [20] | Hideg É, Jansen MAK, Strid Å (2013) UVB exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci 18: 107-115. https://doi.org/10.1016/j.tplants.2012.09.003 |
| [21] |
Köhler H, Contreras RA, Pizarro M, et al. (2017) Antioxidant responses induced by UV-B radiation in deschampsia antarctica Desv. Front Plant Sci 8: 921. https://doi.org/10.3389/fpls.2017.00921
|
| [22] |
Sun M, Gu X, Fu H, et al. (2010) Change of secondary metabolites in leaves of Ginkgo biloba L. in response to UV-B induction. Innov Food Sci Emerg 11: 672-676. https://doi.org/10.1016/j.ifset.2010.08.006
|
| [23] | Dang YF (2015) Influence of enhanced UV-B radiation on Prunella vulgaris ‘growth and various of chemical index components [dissertation], Xi'an: Northwest University. Xi'an: . |
| [24] |
Liu RL, Xiong QJ, Shu Q, et al. (2012) Hyperoside protects cortical neurons from oxygen-glucose deprivation—reperfusion induced injury via nitric oxide signal pathway. Brain Res 1469: 164-173. https://doi.org/10.1016/j.brainres.2012.06.044
|
| [25] | Hameed A, Shah TB, Atta BM, et al. (2008) Gamma irradiation effects on seed germination and growth, protein content, peroxidase and protease activity, lipid peroxidation in desi and kabuli chickpea. Pak J Bot 40: 1033-1041. |
| [26] |
Harrison K, Were LM (2007) Effect of gamma irradiation on total phenolic content yield and antioxidant capacity of almond skin extracts. Food Chem 102: 932-937. https://doi.org/10.1016/j.foodchem.2006.06.034
|




Figures(5) / Tables(1)

Reham Ebrahim, Aya Abdelrazek, Hamed El-Shora, Abu Bakr El-Bediwi. Effect of ultraviolet radiation on molecular structure and photochemical compounds of Salvia hispanica medical seeds[J]. AIMS Biophysics, 2022, 9(2): 172-181. doi: 10.3934/biophy.2022015







 DownLoad:
DownLoad: