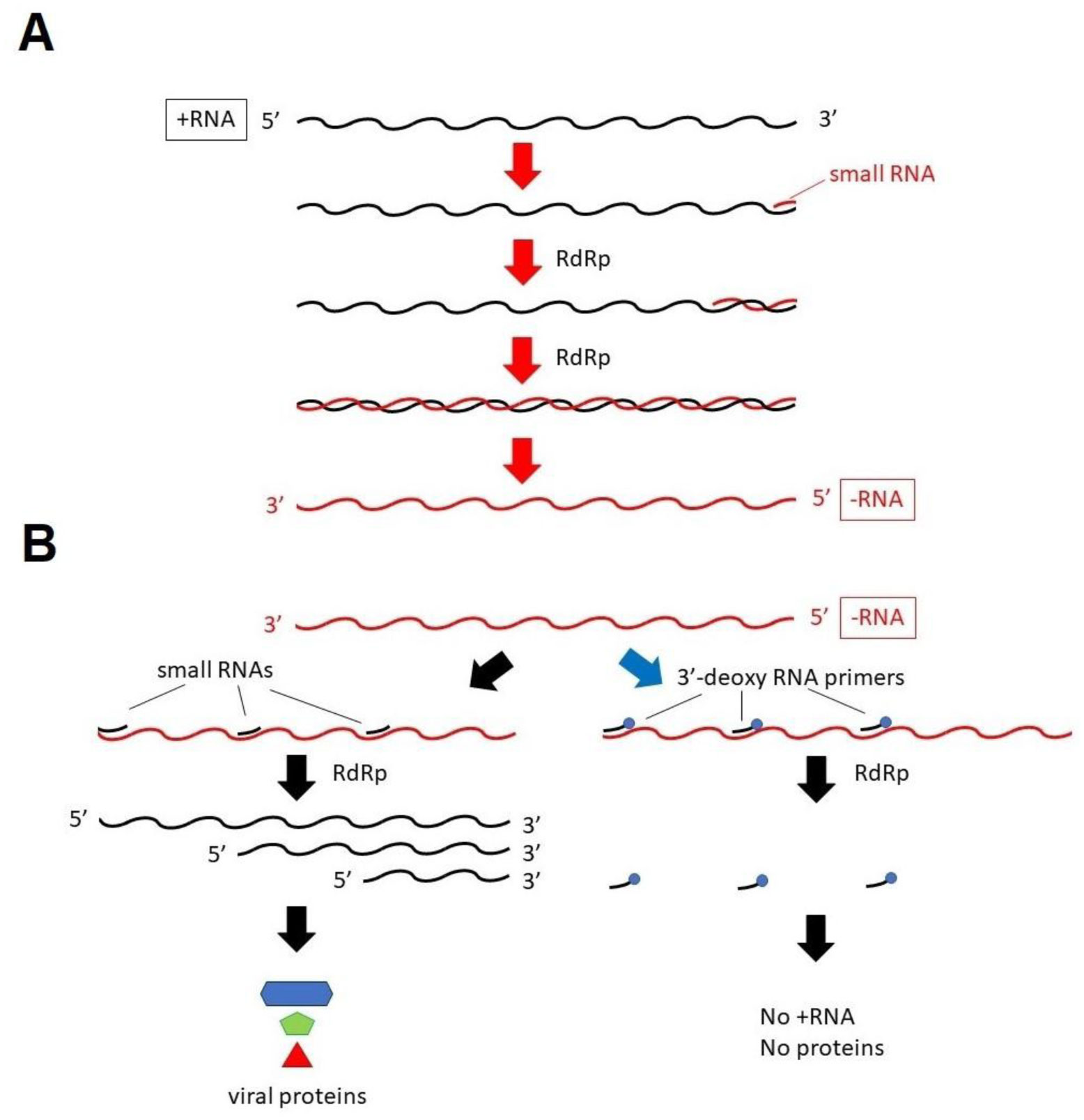
The life cycle of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which is the etiologic agent for coronavirus disease-19 (COVID-19), has been revealed, and the molecular mechanism of how this virus replicates has been studied. Effective drugs could be designed to target one of the replication processes. This virus, which contains plus-strand RNA as a genome, is characteristic because it synthesizes its genome and other transcripts with minus-strand template RNA. These viral RNAs, including its genome, are transcribed by an RNA-dependent RNA polymerase (RdRp). In this article, we discuss whether novel materials from RNA derivatives could be applied to COVID-19 patients to prevent or disturb viral transcription.
Citation: Fumiaki Uchiumi, Yoko Ogino, Akira Sato, Sei-ichi Tanuma. Insights into the development of effective materials to suppress replication of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)[J]. AIMS Bioengineering, 2020, 7(3): 124-129. doi: 10.3934/bioeng.2020012
The life cycle of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which is the etiologic agent for coronavirus disease-19 (COVID-19), has been revealed, and the molecular mechanism of how this virus replicates has been studied. Effective drugs could be designed to target one of the replication processes. This virus, which contains plus-strand RNA as a genome, is characteristic because it synthesizes its genome and other transcripts with minus-strand template RNA. These viral RNAs, including its genome, are transcribed by an RNA-dependent RNA polymerase (RdRp). In this article, we discuss whether novel materials from RNA derivatives could be applied to COVID-19 patients to prevent or disturb viral transcription.

| [1] | Carey MF, Peterson CL, Smale ST (2009) A primer on transcriptional regulation in mammalian cells. Transcriptional Regulation in Eukaryotes: Concepts, Strategies, and Techniques NY: Cold Spring Harbor Laboratory Press, 1-45. |
| [2] |
Chen Y, Liu Q, Guo D (2020) Emerging coronaviruses: genome structure, replication, and pathogenesis. J Med Virol 92: 418-423. doi: 10.1002/jmv.25681

|
| [3] |
Wu F, Zhao S, Yu B, et al. (2020) A new coronavirus associated with human respiratory disease in China. Nature 579: 265-269. doi: 10.1038/s41586-020-2008-3
|
| [4] |
Te Velthuis AJW, Arnold JJ, Cameron CE, et al. (2010) The RNA polymerase activity of SARS-coronavirus nsp12 is primer dependent. Nucleic Acids Res 38: 203-214. doi: 10.1093/nar/gkp904
|
| [5] |
Ahn DG, Chi JK, Taylor DR, et al. (2012) Biochemical characterization of a recombinant SARS coronavirus nsp12 RNA-dependent RNA polymerase capable of copying viral RNA templates. Arch Virol 157: 2095-2104. doi: 10.1007/s00705-012-1404-x
|
| [6] |
Lung J, Lin YS, Yang YH, et al. (2020) The potential chemical structure of anti-SARS-CoV-2 RNA-dependent RNA polymerase. J Med Virol 92: 693-697. doi: 10.1002/jmv.25761
|
| [7] |
Tan YW, Fung TS, Shen H, et al. (2018) Coronavirus infectious bronchitis virus non-structural proteins 8 and 12 form stable complex independent of the non-translated regions of viral RNA and other viral proteins. Virology 513: 75-84. doi: 10.1016/j.virol.2017.10.004
|
| [8] | Joyce GF, Orgel LE (2006) Progress toward understanding the origin of the RNA world. The RNA World NY: Cold Spring Harbor Laboratory Press, 23-56. |
| [9] | Cech TR (2011) The RNA world in context. RNA Worlds: From Life's Origins to Diversity in Gene Regulation NY: Cold Spring Harbor Laboratory Press, 1-5. |
| [10] |
Snijder EJ, Bredenbeek PJ, Dobbe JC, et al. (2003) Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage. J Mol Biol 331: 991-1004. doi: 10.1016/S0022-2836(03)00865-9
|
| [11] | Song T, Ma K, Zhao C, et al. (2019) MicroRNA-2053 overexpression inhibits the development and progression of hepatocellular carcinoma. Oncol Lett 18: 2043-2049. |
| [12] |
Chun N, Coca SG, He JC (2018) A protective role for microRNA-668 in acute kidney injury. J Clin Invest 128: 5216-5218. doi: 10.1172/JCI124923
|
| [13] |
Wei Q, Sun H, Song S, et al. (2018) MicroRNA-668 represses MTP18 to preserve mitochondrial dynamics in ischemic acute kidney injury. J Clin Invest 128: 5448-5464. doi: 10.1172/JCI121859
|
| [14] |
Trobaugh DW, Klimstra WB (2017) MicroRNA regulation of RNA virus replication and pathogenesis. Trends Mol Med 23: 80-93. doi: 10.1016/j.molmed.2016.11.003
|
| [15] |
Liu K, Chen Y, Lin R, et al. (2020) Clinical features of COVID-19 in elderly patients: a comparison with young and middle-aged patients. J Infect 80: e14-e18. doi: 10.1016/j.jinf.2020.03.005
|
| [16] |
Zagryazhskaya A, Zhivotovsky B (2014) miRNA in lung cancer: a link to aging. Ageing Res Rev 17: 54-67. doi: 10.1016/j.arr.2014.02.009
|
| [17] |
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74: 5463-5467. doi: 10.1073/pnas.74.12.5463
|
| [18] |
Nakayama C, Saneyoshi M (1985) Differential inhibitory effects of 5-substituted 1-β-d-xylofuranosyluracil 5′-triphosphates and related nucleotides on DNA-dependent RNA polymerases I and II from the cherry salmon (Oncorhynchus masou). J Biochem 98: 417-425. doi: 10.1093/oxfordjournals.jbchem.a135296
|
| [19] |
Furuta Y, Komeno T, Nakamura T (2017) Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase. Proc Jpn Acad Ser B 93: 449-463. doi: 10.2183/pjab.93.027
|
| [20] |
Gampe C, White ACS, Siva S, et al. (2018) 3′-Modification stabilizes mRNA and increases translation in cells. Bioorg Med Chem Lett 28: 2451-2453. doi: 10.1016/j.bmcl.2018.06.008
|
| [21] |
Tan CW, Chan YF, Quah YW, et al. (2014) Inhibition of enterovirus 71 infection by antisense octaguanidinium dendrimer-conjugated morpholino oligomers. Antivir Res 107: 35-41. doi: 10.1016/j.antiviral.2014.04.004
|
| [22] |
Chung TD, Cianci C, Hagen M, et al. (1994) Biochemical studies on capped RNA primers identify a class of oligonucleotide inhibitors of the influenza virus RNA polymerase. Proc Natl Acad Sci USA 91: 2372-2376. doi: 10.1073/pnas.91.6.2372
|
| [23] |
Subissi L, Imbert I, Ferron F, et al. (2014) SARS-CoV ORF1b-encoded nonstructural proteins 12–16: Replicative enzymes as antiviral targets. Antivir Res 101: 122-130. doi: 10.1016/j.antiviral.2013.11.006
|
| [24] |
Fujita Y, Kuwano K, Ochiya T (2015) Development of small RNA delivery systems for lung cancer therapy. Int J Mol Sci 16: 5254-5270. doi: 10.3390/ijms16035254
|
| [25] |
Walsh C (2000) Molecular mechanisms that confer antibacterial drug resistance. Nature 406: 775-781. doi: 10.1038/35021219
|
| [26] |
Van Aerschot A, Herdewijn P, Janssen G, et al. (1989) Synthesis and antiviral activity evaluation of 3′-fluoro-3′-deoxyribonucleosides: broad-spectrum antiviral activity of 3′-fluoro-3′-deoxyadenosine. Antivir Res 12: 133-150. doi: 10.1016/0166-3542(89)90047-8
|
| [27] | Uchiumi F (2018) Introductory chapter: Current studies in transcriptional control system; toward the establishment of therapies against human diseases. Gene Expression and Regulation in Mammalian Cells: Transcription From General Aspects London: InTech OPEN, 3-13. |
| [28] | Uchiumi F, Asai M (2018) Gene expression controlling system and its application to medical sciences. Gene Expression and Control London: InTech OPEN, 3-15. |
Figures(1)

Fumiaki Uchiumi, Yoko Ogino, Akira Sato, Sei-ichi Tanuma. Insights into the development of effective materials to suppress replication of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)[J]. AIMS Bioengineering, 2020, 7(3): 124-129. doi: 10.3934/bioeng.2020012







 DownLoad:
DownLoad: