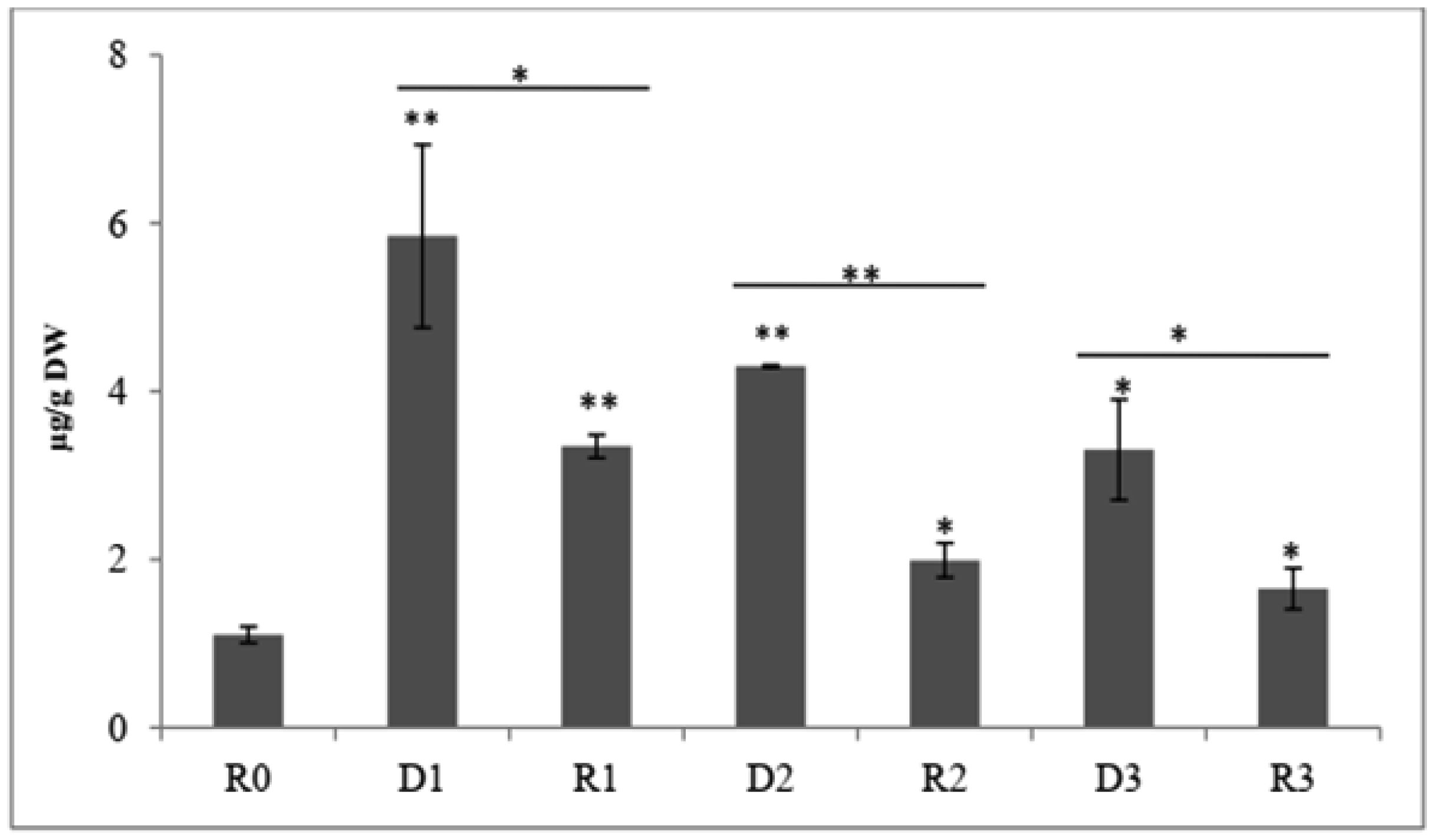
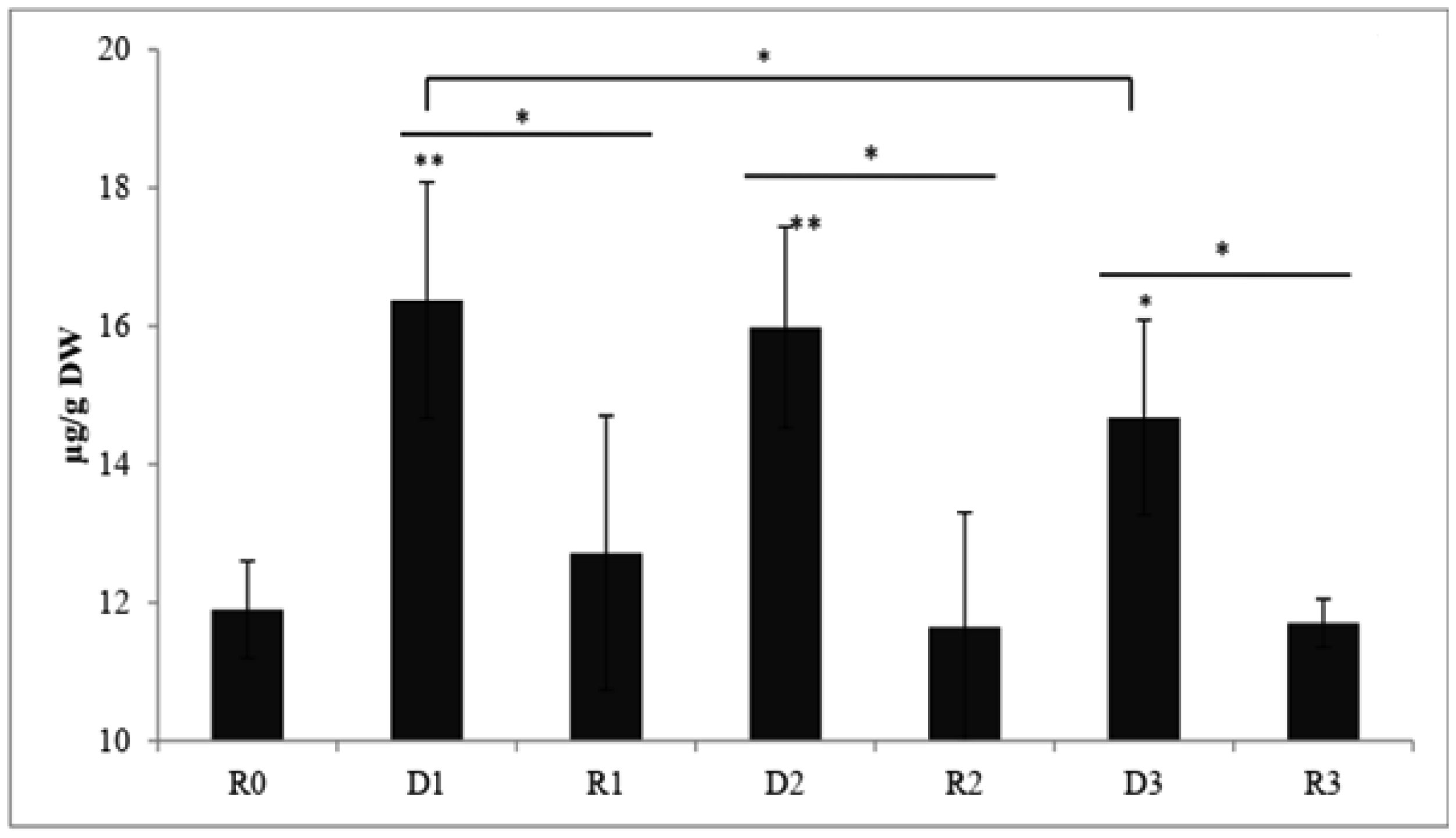
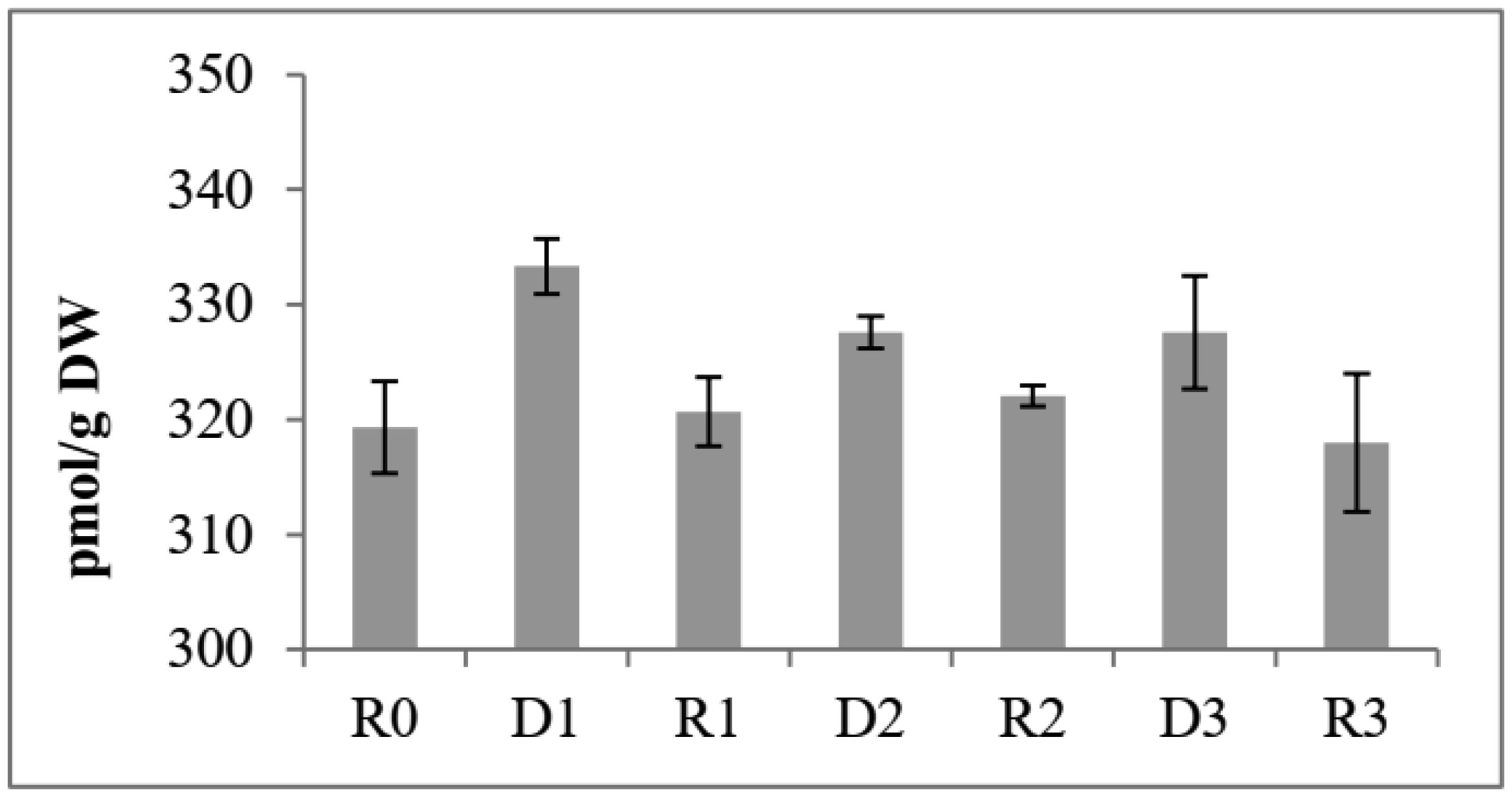
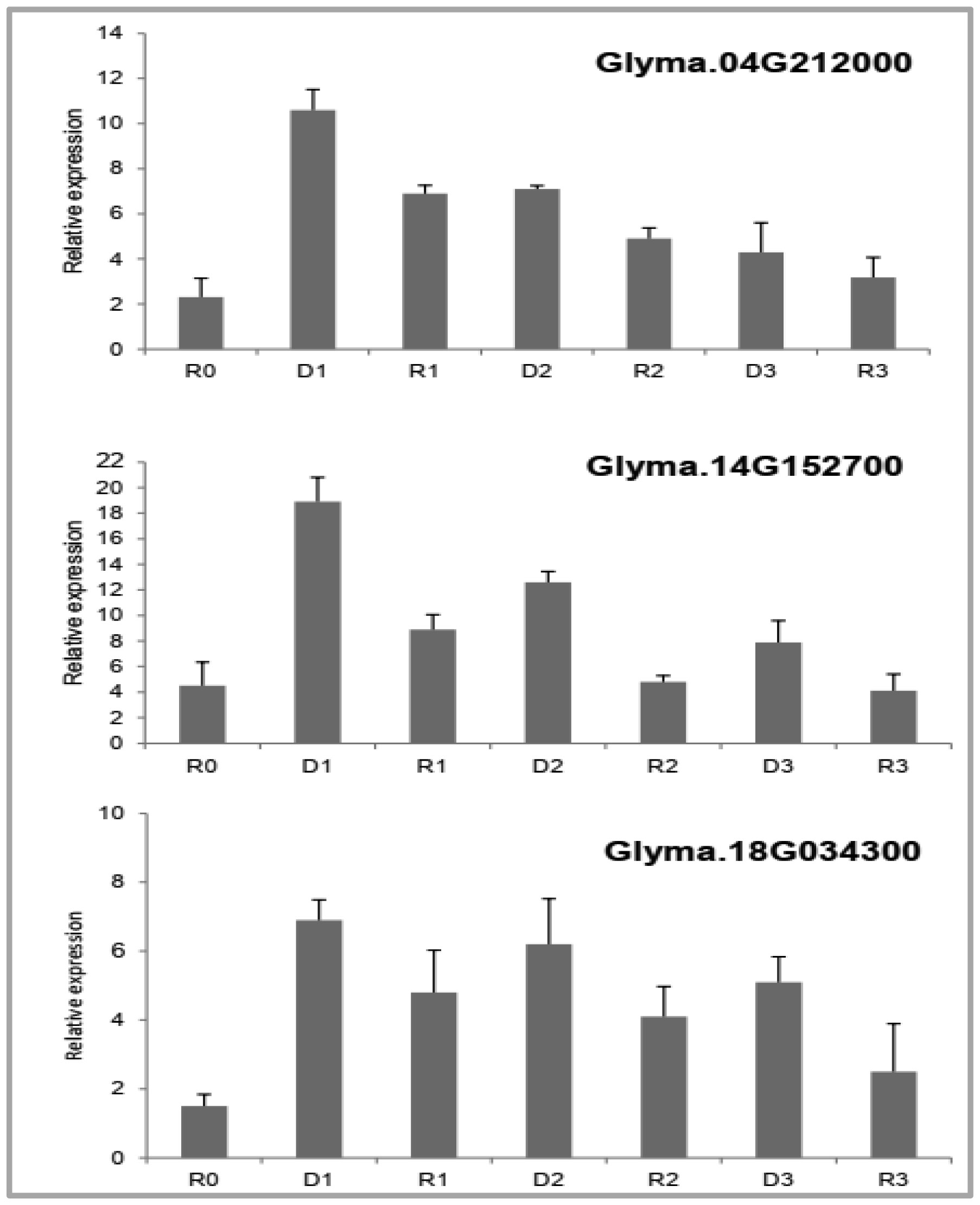
Drought stress memory in plant can alter their physiological, biochemical and molecular to a subsequent stress. An experiment was conducted to determine biochemical parameters of soybean seedlings under drought stress memory. 14-days-old soybean seedlings were subjected to three consecutive water deficit phases (D1, D2, D3), each phase recovered by re-watering (R1, R2, R3), and control plant watering daily (R0). Leave of seedlings from these phases were collected and analyzed. Significantly increasing contents of soluble sugar and proline observed at the first drought stress. After the first re-water and continuous stress phases, these indicator contents decreased and maintained at a relatively stable level. Expression level of two memory genes encoded to transcription factor (NAC09 and NAC109) and one gene encoded enzyme Δ1-pyrroline-5-carboxylate synthetase (P5CS1) were increased in the first stress and decreased in third stress. Our results demonstrate that changing of biochemical parameters of soybean seedlings can be seen as the strong indications of “drought stress memory”. This result may serve as a reference platform to study advanced researches at molecular and genetic levels.
Citation: Thi Thuy Quynh Nguyen, Le Thanh Huyen Trinh, Hoang Bao Vy Pham, Tri Vien Le, Thi Kim Hue Phung, Suk-Ha Lee, Jong-Joo Cheong. Evaluation of proline, soluble sugar and ABA content in soybean Glycine max (L.) under drought stress memory[J]. AIMS Bioengineering, 2020, 7(3): 114-123. doi: 10.3934/bioeng.2020011
Drought stress memory in plant can alter their physiological, biochemical and molecular to a subsequent stress. An experiment was conducted to determine biochemical parameters of soybean seedlings under drought stress memory. 14-days-old soybean seedlings were subjected to three consecutive water deficit phases (D1, D2, D3), each phase recovered by re-watering (R1, R2, R3), and control plant watering daily (R0). Leave of seedlings from these phases were collected and analyzed. Significantly increasing contents of soluble sugar and proline observed at the first drought stress. After the first re-water and continuous stress phases, these indicator contents decreased and maintained at a relatively stable level. Expression level of two memory genes encoded to transcription factor (NAC09 and NAC109) and one gene encoded enzyme Δ1-pyrroline-5-carboxylate synthetase (P5CS1) were increased in the first stress and decreased in third stress. Our results demonstrate that changing of biochemical parameters of soybean seedlings can be seen as the strong indications of “drought stress memory”. This result may serve as a reference platform to study advanced researches at molecular and genetic levels.

| [1] |
Li X, Cai J, Liu F, et al. (2014) Cold priming drives the sub-cellular antioxidant systems to protect photosynthetic electron transport against subsequent low temperature stress in winter wheat. Plant Physiol Biochem 82: 34-43. doi: 10.1016/j.plaphy.2014.05.005

|
| [2] |
Muscolo A, Junker A, Klukas C, et al. (2015) Phenotypic and metabolic responses to drought and salinity of four contrasting lentil accessions. J Exp Bot 66: 5467-5480. doi: 10.1093/jxb/erv208
|
| [3] |
Saxena SC, Kaur H, Verma P, et al. (2013) Osmoprotectants: potential for crop improvement under adverse conditions. Plant Acclimation to Environmental Stress New York: Springer, 197-232. doi: 10.1007/978-1-4614-5001-6_9
|
| [4] |
Meena M, Divyanshu K, Kumar S, et al. (2019) Regulation of L-proline biosynthesis, signal transduction, transport, accumulation, and its vital role in plants during variable environmental conditions. Heliyon 5: e02952. doi: 10.1016/j.heliyon.2019.e02952
|
| [5] |
Feng XJ, Li JR, Qi SL, et al. (2016) Light affects salt stress-induced transcriptional memory of P5CS1 in Arabidopsis. Proc Natl Acad Sci USA 113: E8335-E8343. doi: 10.1073/pnas.1610670114
|
| [6] |
Székely G, Ábrahám E, Cséplő Á, et al. (2008) Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J 53: 11-28. doi: 10.1111/j.1365-313X.2007.03318.x
|
| [7] |
Sharma A, Shahzad B, Kumar V, et al. (2019) Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 9: 285. doi: 10.3390/biom9070285
|
| [8] |
Le DT, Nishiyama RIE, Watanabe Y, et al. (2011) Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res 18: 263-276. doi: 10.1093/dnares/dsr015
|
| [9] |
Yang X, Wang X, Ji L, et al. (2015) Overexpression of a Miscanthus lutarioriparius NAC gene MlNAC5 confers enhanced drought and cold tolerance in Arabidopsis. Plant cell Rep 34: 943-958. doi: 10.1007/s00299-015-1756-2
|
| [10] |
Kinoshita T, Seki M (2014) Epigenetic memory for stress response and adaptation in plants. Plant Cell Physiol 55: 1859-1863. doi: 10.1093/pcp/pcu125
|
| [11] |
Wang X, Vignjevic M, Jiang D, et al. (2014) Improved tolerance to drought stress after anthesis due to priming before anthesis in wheat (Triticum aestivum L.) var. Vinjett. J Exp Bot 65: 6441-6456. doi: 10.1093/jxb/eru362
|
| [12] |
Avramova Z (2015) Transcriptional ‘memory’ of a stress; transient chromatin and memory (epigenetic) marks at stress-response genes. Plant J 83: 149-159. doi: 10.1111/tpj.12832
|
| [13] |
Beckers GJM, Conrath U (2007) Priming for stress resistance: from the lab to the field. Curr Opin Plant Biol 10: 425-431. doi: 10.1016/j.pbi.2007.06.002
|
| [14] |
Ramírez DA, Rolando JL, Yactayo W, et al. (2015) Improving potato drought tolerance through the induction of long-term water stress memory. Plant Sci 238: 26-32. doi: 10.1016/j.plantsci.2015.05.016
|
| [15] |
Sani E, Herzyk P, Perrella G, et al. (2013) Hyperosmotic priming of Arabidopsis seedlings establishes a long-term somatic memory accompanied by specific changes of the epigenome. Genome Biol 14: R59. doi: 10.1186/gb-2013-14-6-r59
|
| [16] |
Wehner G, Balko C, Ordon F (2016) Experimental design to determine drought stress response and early leaf senescence in barley (Hordeum vulgare L.). Bio-Protocol 6: 1-16. doi: 10.21769/BioProtoc.1749
|
| [17] |
Sharma S, Villamor JG, Verslues PE (2011) Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol 157: 292-304. doi: 10.1104/pp.111.183210
|
| [18] |
Leufen G, Noga G, Hunsche M (2016) Drought stress memory in sugar beet: mismatch between biochemical and physiological parameters. J Plant Growth Regul 35: 680-689. doi: 10.1007/s00344-016-9571-8
|
| [19] |
Arabzadeh N (2012) The effect of drought stress on soluble carbohydrates (sugars) in two species of Haloxylon persicum and Haloxylon aphyllum. Asian J Plant Sci 11: 44-51. doi: 10.3923/ajps.2012.44.51
|
| [20] | Da Silva Lobato AK, De Oliveira Neto CF, Dos Santos Filho BG, et al. (2008) Physiological and biochemical behavior in soybean (Glycine max cv. Sambaiba) plants under water deficit. Aust J Crop Sci 2: 25-32. |
| [21] |
Li TH, Li SH (2005) Leaf responses of micropopagated apple plants to water stress: nonstructural carbohydrate composition and regulatory role of metabolic enzymes. Tree Physiol 25: 495-504. doi: 10.1093/treephys/25.4.495
|
| [22] |
Li P, Yang H, Wang L, et al. (2019) Physiological and transcriptome analyses reveal short-term responses and formation of memory under drought stress in rice. Front Genet 10: 55. doi: 10.3389/fgene.2019.00055
|
| [23] |
Liu J, Jiang MY, Zhou YF, et al. (2005) Production of polyamines is enhanced by endogenous abscisic acid in maize seedlings subjected to salt stress. J Integr Plant Biol 47: 1326-1334. doi: 10.1111/j.1744-7909.2005.00183.x
|
| [24] | Faltusová-Kadlecová Z, Faltus M, Prášil I (2002) Abscisic acid content during cold hardening of barley and wheat cultivars with different freezing tolerance. Rostl Výroba 48: 490-493. |
| [25] |
Iovieno P, Punzo P, Guida G, et al. (2016) Transcriptomic changes drive physiological responses to progressive drought stress and rehydration in tomato. Front Plant Sci 7: 371. doi: 10.3389/fpls.2016.00371
|
| [26] |
Huang X, Chen MH, Yang LT, et al. (2015) Effects of exogenous abscisic acid on cell membrane and endogenous hormone contents in leaves of sugarcane seedlings under cold stress. Sugar Tech 17: 59-64. doi: 10.1007/s12355-014-0343-0
|
| [27] |
Ding Y, Virlouvet L, Liu N, et al. (2014) Dehydration stress memory genes of Zea mays; comparison with Arabidopsis thaliana. BMC Plant Biol 14: 141. doi: 10.1186/1471-2229-14-141
|
| [28] |
Fujita Y, Fujita M, Shinozaki K, et al. (2011) ABA-mediated transcriptional regulation in response to osmotic stress in plants. J Plant Res 124: 509-525. doi: 10.1007/s10265-011-0412-3
|
| [29] |
Jiang H, Li H, Bu Q, et al. (2009) The RHA2a-interacting proteins ANAC019 and ANAC055 may play a dual role in regulating ABA response and jasmonate response. Plant Signal Behav 4: 464-466. doi: 10.4161/psb.4.5.8543
|
| [30] |
Khan R, Zhou P, Ma X, et al. (2019) Transcriptome profiling, biochemical and physiological analyses provide new insights towards drought tolerance in Nicotiana tabacum L. Genes 10: 1041. doi: 10.3390/genes10121041
|
Figures(4) / Tables(1)

Thi Thuy Quynh Nguyen, Le Thanh Huyen Trinh, Hoang Bao Vy Pham, Tri Vien Le, Thi Kim Hue Phung, Suk-Ha Lee, Jong-Joo Cheong. Evaluation of proline, soluble sugar and ABA content in soybean Glycine max (L.) under drought stress memory[J]. AIMS Bioengineering, 2020, 7(3): 114-123. doi: 10.3934/bioeng.2020011







 DownLoad:
DownLoad: