Citation: Larisa M. Somova, Boris G. Andryukov, Irina N. Lyapun. Cell geteromorphism in the conditions of persistence of sapronoses causative agents in various environments[J]. AIMS Microbiology, 2019, 5(2): 147-157. doi: 10.3934/microbiol.2019.2.147

| [1] | Bukharin OV, Gunzburg AP, Romanova YuM, et al. (2005) Mechanisms of bacteria survival. M: Medicine 365. |
| [2] | Litvin VYu, Gunzburg AL, Pushkareva VI, et al. (1998) Epidemiological aspects of the bacteria ecology. M: Farmaus-Print 255. |
| [3] | Litvin VYu, Somov GP, Pushkareva VI (2010) Sapronoses as a natural focal disease. Epidemiol Vaccinoprophyl 50: 10–16. |
| [4] | Somov GP (2001) The modern idea about sapronoses (the main results of the study of the problem). Pacific Med J 2: 67–70. |
| [5] | Bukharin OV (2015) Infectious symbiology. J Microbiol 4: 4–9. |
| [6] | Somov GP, Buzoleva LS (2004) Adaptation of pathogenic bacteria to abiotic environmental factors. Vladivostok: Primpigraph Combine 167. |
| [7] |
Ayrapetyan M, Williams TC, Oliver JD (2014) Interspecific quorum sensing mediates the resuscitation of viable but non-culturable vibrios. Appl Environ Microbiol 80: 2478–2483. doi: 10.1128/AEM.00080-14

|
| [8] |
Boaretti M, Lleo MM, Bonato B, et al. (2003) Involvement of rpoS in the survival of Escherichia coli in the viable but non-culturable state. Environ Microbiol 5: 986–996. doi: 10.1046/j.1462-2920.2003.00497.x
|
| [9] |
Hong SH, Wang XX, O'Connor HF, et al. (2012) Bacterial persistence increases as environmental fitness decreases. Microb Biotechnol 5: 509–522. doi: 10.1111/j.1751-7915.2011.00327.x
|
| [10] |
Kuris AM, Lafferty KD, Sokolow SH (2014) Sapronosis: a distinctive type of infectious agent. Trends Parasitol 30: 386–393. doi: 10.1016/j.pt.2014.06.006
|
| [11] | Milko ES, Egorov NS (1991) Heterogeneity of the bacterial population and the process of dissociation. M: Moscow State University 142. |
| [12] |
Ayrapetyan M, Williams TC, Baxter R, et al. (2015) Viable but non-culturable and persister cells coexist stochastically and are induced by human serum. Infect Immun 83: 4194–4203. doi: 10.1128/IAI.00404-15
|
| [13] |
Baffone W, Citterio B, Vittoria E, et al. (2003) Retention of virulence in viable but non-culturable halophilic Vibrio spp. Int J Food Microbiol 89: 31–39. doi: 10.1016/S0168-1605(03)00102-8
|
| [14] | Kim J-S, Chowdhury N, Wood TK (2017) Viable but non-culturable cells are persister cells. Environ Microbiol 20: 2038–2048. |
| [15] | Li L, Mendis N, Trigui H, et al. (2014) The importance of the viable but nonculturable state in human bacterial pathogens. Front Microbiol 5: 258. |
| [16] | Isachkova LM, Zhavoronkov AA, Shubin FN (1993) L-transformation of Yersinia in experimental pseudotuberculosis. J Microbiol Epidemiol Immunobiol 1: 11–15. |
| [17] |
Joseleau-Petit D, Liébart JC, Ayala JA, et al. (2007) Unstable Escherichia coli L-forms revisited: Growth requires peptidoglycan synthesis. J Bacteriol 189: 6512–6520. doi: 10.1128/JB.00273-07
|
| [18] |
Hayes F (2003) Toxins-antitoxins: Plasmid maintenance, programmed cell death, and cell cycle arrest. Science 301: 1496–1499. doi: 10.1126/science.1088157
|
| [19] |
Page R, Peti W (2016) Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat Chem Biol 12: 208–214. doi: 10.1038/nchembio.2044
|
| [20] |
Wood TK (2016) Combatting bacterial persister cells. Biotechnol Bioeng 113: 476–83. doi: 10.1002/bit.25721
|
| [21] | Ayrapetyan M, Williams TC, Oliver JD (2015) Bridging the gap between viable but non-culturable and antibiotic persistent bacteria. Trends Microbiol 23: 7–13. |
| [22] |
Nelson EJ, Chowdhury A, Flynn J, et al. (2008) Transmission of Vibrio cholerae is antagonized by lytic phage and entry into the aquatic environment. PLoS Pathog 4: e1000187. doi: 10.1371/journal.ppat.1000187
|
| [23] |
Nowakowska J, Oliver JD (2013) Resistance to environmental stresses by Vibrio vulnificus in the viable but nonculturable state. FEMS Microbiol Ecol 84: 213–222. doi: 10.1111/1574-6941.12052
|
| [24] | Belov AB, Kulikalova ES (2016) Sapronoses: ecology of pathogens, epidemiology and taxonomy. Epidemiol Vaccinoprophyl 86: 5–16. |
| [25] | Brusina EB (2015) Epidemiology of infections associated with the provision of medical care, caused by pathogens of the sapronosis group. Epidemiol Vaccinoprophyl 81: 50–56. |
| [26] | Pushkareva VI, Litvin VYu, Konstantinova ND (1990) Analysis of the mechanisms of interactions of Yersinia with infusoria Tetrahymena pyriformis at the cellular and subcellular levels. J Microbiol Epidemiol Immunobiol 1: 3–8. |
| [27] | Kirillova FM, Timchenko NF (1984) Electron-microscopic study of the interaction of Yersinia pseudotuberculosis with macrophages and HeLa cells. J Microbiol Epidemiol Immunobiol 7: C 95–97. |
| [28] | Somova LM, Buzoleva LS, Plekhova NG (2009) Ultrastructure of pathogenic bacteria in different environmental conditions. Vladivostok: Medicine DV 199. |
| [29] | Belov AB, Kuzin AA (2017) Sapronoses infections associated with the provision of medical care: problematic issues of the theory of epidemiology. Permsky Medical J 4: 94–102. |
| [30] | Amara AA, Salem-Bekhit MM, Alanazi FK (2013) Sponge-like: a new protocol for preparing bacterial ghosts. Sci World J 2013: 545741. |
| [31] | Oliver JD (2005) The viable but nonculturable state in bacteria. J Microbiol 43: 93–100. |
| [32] |
Orman MA, Brynildsen MP (2013) Establishment of a method to rapidly assay bacterial persister metabolism. Antimicrob Agents Ch 57: 4398–4409. doi: 10.1128/AAC.00372-13
|
| [33] |
Lennon JT, Jones SE (2011) Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat Rev Microbiol 9: 119–130. doi: 10.1038/nrmicro2504
|
| [34] | Requena JM (2012) Stress response in microbiology. Horizon Scientific Press, 436. |
| [35] | Storz G, Hengge R (2010) Bacterial stress responses. American Society for Microbiology : ASM Press, 26. |
| [36] |
Oliver JD (2010) Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol Rev 34: 415–425. doi: 10.1111/j.1574-6976.2009.00200.x
|
| [37] |
Hobby GL, Meyer K, Chaffee E (1942) Observations on the mechanism of action of penicillin. Exp Biol Med 50: 281–285. doi: 10.3181/00379727-50-13773
|
| [38] | Goncalves FD, de Carvalho CC (2016) Phenotypic modifications in Staphylococcus aureus cells exposed to high concentrations of vancomycin and teicoplanin. Front Microbiol 7: 13. |
| [39] |
Korch SB, Henderson TA, Hill TM (2003) Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol Microbiol 50: 1199–1213. doi: 10.1046/j.1365-2958.2003.03779.x
|
| [40] |
Mulcahy LR, Burns JL, Lory S, et al. (2010) Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J Bacteriol 192: 6191–6199. doi: 10.1128/JB.01651-09
|
| [41] |
Rivers B, Steck TR (2001) Viable but non-culturable uropathogenic bacteria are present in the mouse urinary tract following urinary tract infection and antibiotic therapy. Urol Res 29: 60–66. doi: 10.1007/s002400000151
|
| [42] | Colwell RR (2009) Viable but Not Cultivable Bacteria, In: Uncultivated Microorganisms, 1 Eds, Springer-Verlag Berlin Heidelberg, 121–129. |
| [43] |
Kusumoto A, Asakura H, Kawamoto K (2012) General stress sigma factor RpoS influences time required to enter the viable but non-culturable state in Salmonella Enterica. Microbiol Immunol 56: 228–237. doi: 10.1111/j.1348-0421.2012.00428.x
|
| [44] |
Helaine S, Kugelberg E (2014) Bacterial persisters: formation, eradication, and experimental systems. Trends Microbiol 22: 417–424. doi: 10.1016/j.tim.2014.03.008
|
| [45] |
Potgieter M, Bester J, Kell DB, et al. (2015) The dormant blood microbiome in chronic, inflammatory diseases. FEMS Microbiol Rev 39: 567–591. doi: 10.1093/femsre/fuv013
|
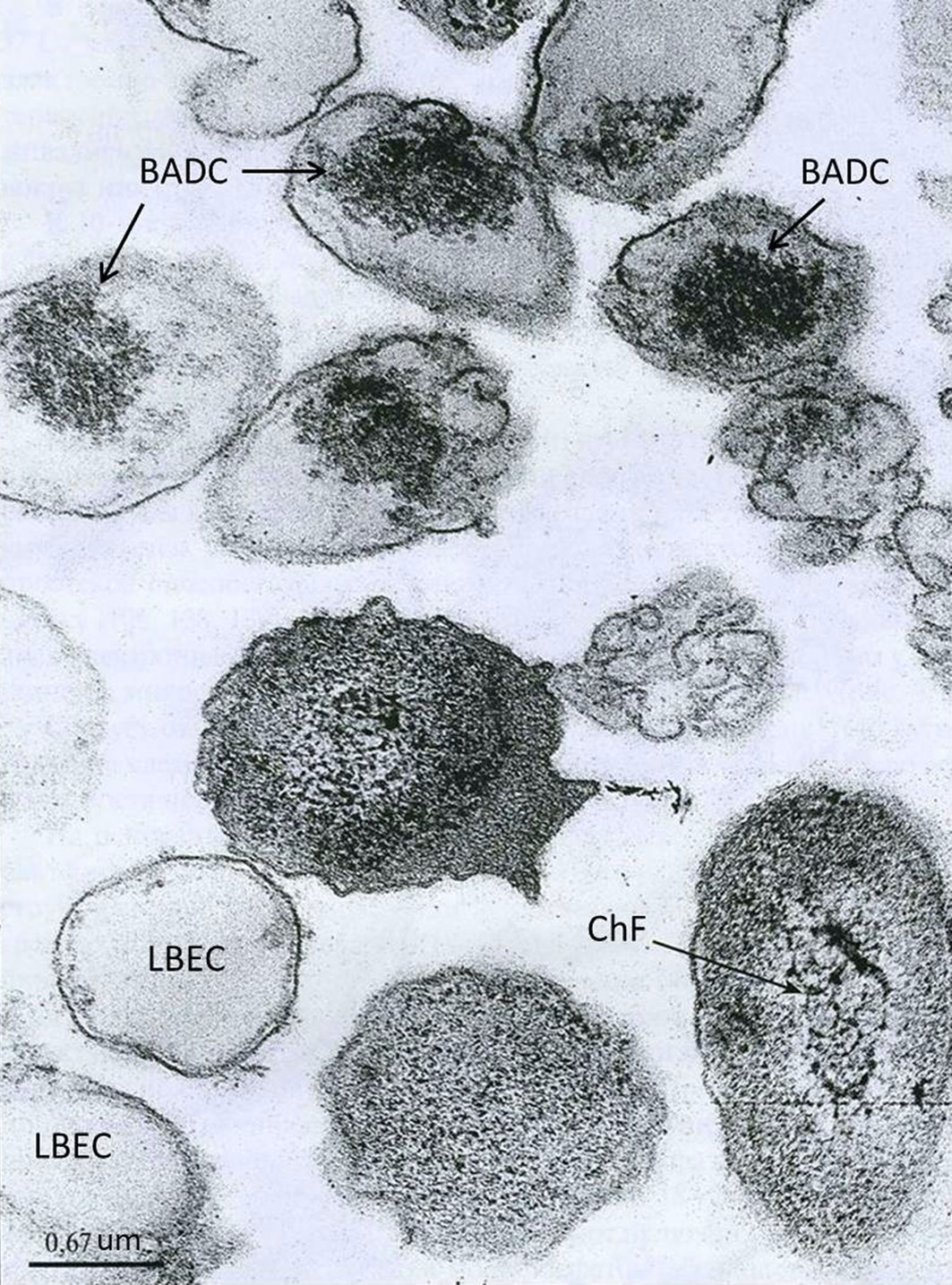
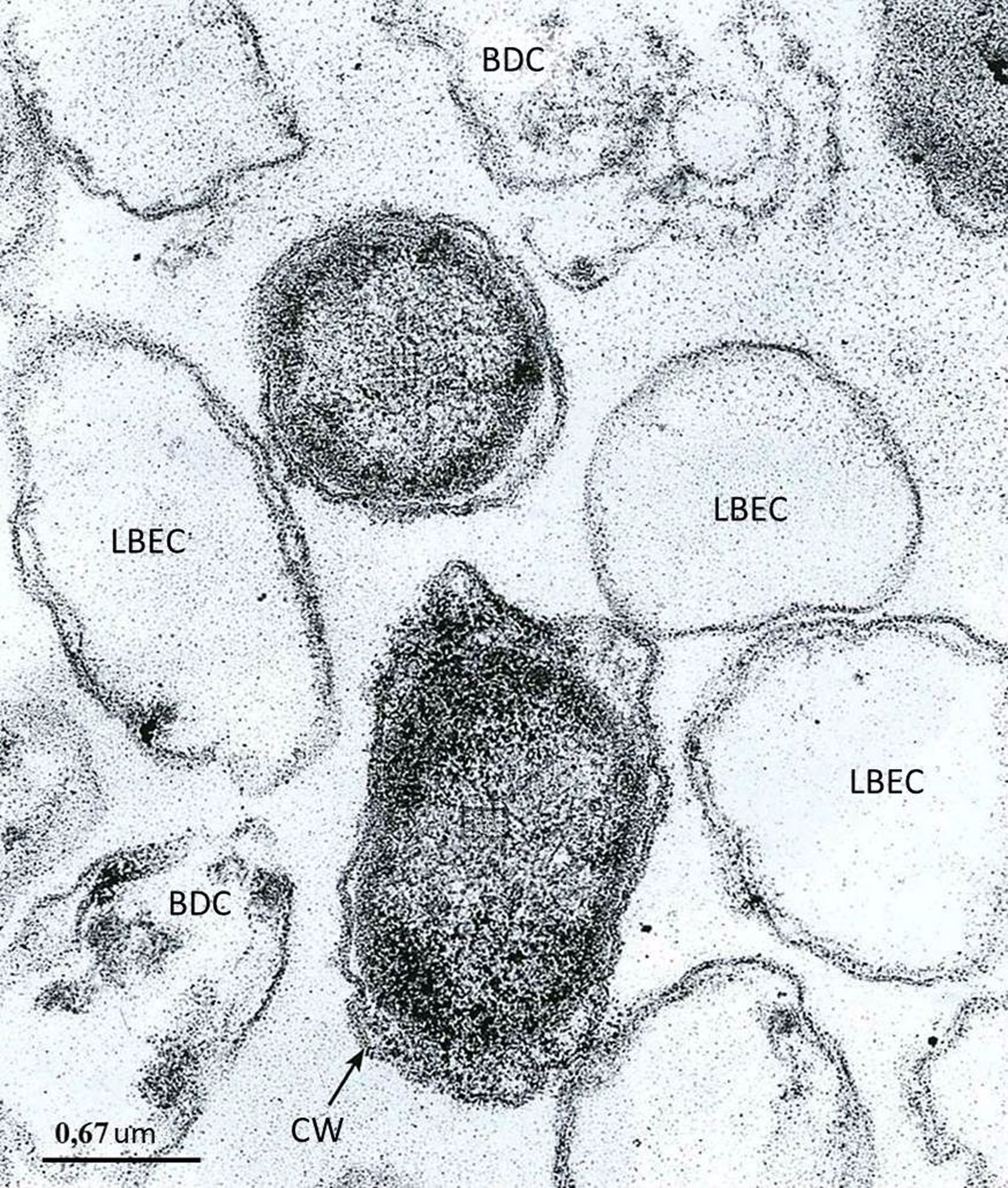
Figures(2)

Larisa M. Somova, Boris G. Andryukov, Irina N. Lyapun. Cell geteromorphism in the conditions of persistence of sapronoses causative agents in various environments[J]. AIMS Microbiology, 2019, 5(2): 147-157. doi: 10.3934/microbiol.2019.2.147







 DownLoad:
DownLoad: