Citation: Tuan Syaripah Najihah, Mohd Hafiz Ibrahim, Nurul Amalina Mohd Zain, Rosimah Nulit, Puteri Edaroyati Megat Wahab. Activity of the oil palm seedlings exposed to a different rate of potassium fertilizer under water stress condition[J]. AIMS Environmental Science, 2020, 7(1): 46-68. doi: 10.3934/environsci.2020004

| [1] |
Carr MKV (2011) The water relations and irrigation requirements of oil palm (Elaeis guineensis): A review. Exp Agric 47: 629-652. doi: 10.1017/S0014479711000494

|
| [2] | Kushairi A, Loh SK, Azman I, et al. (2018) Oil palm economic performance in Malaysia and r&d progress in 2017. J Oil Palm Res 30: 163-195. |
| [3] |
Paterson RRM, Lima N (2018) Climate change affecting oil palm agronomy, and oil palm cultivation increasing climate change, require amelioration. Eco Evolut 8: 452-461. doi: 10.1002/ece3.3610
|
| [4] | Paterson RRM, Kumar L, Taylor S, et al. (2015) Future climate effects on suitability for growth of oil palms in Malaysia and Indonesia. Sci R 5: 1-11. |
| [5] | Rosenani AB, Rovica R, Cheah PM, et al. (2016) Growth Performance and Nutrient Uptake of Oil Palm Seedling in Prenursery Stage as Influenced by Oil Palm Waste Compost in Growing Media. Int J Agron. |
| [6] |
Awalludin MF, Sulaiman O, Hashim R, et al. (2015) An overview of the oil palm industry in Malaysia and its waste utilization through thermochemical conversion, specifically via liquefaction. Renew Sustain Energ Rev 50: 1469-1484. doi: 10.1016/j.rser.2015.05.085
|
| [7] | Suresh K (2013) Adaptation and Mitigation Strategies for Climate-Resilient Oil Palm. India. Springer India 199-211. |
| [8] |
Culman M, de Farias CM, Bayona C, et al. (2019) Using agrometeorological data to assist irrigation management in oil palm crops: A decision support method and results from crop model simulation. Agri Water Manage 213: 1047-1062. doi: 10.1016/j.agwat.2018.09.052
|
| [9] | Noor MRM, Harun MH (2004) Importance of water use efficiency (WUE) in oil palm productivity. Oil Palm Bull 2004: 24-30. |
| [10] |
Azzeme AM, Abdullah SNA, Aziz MA, et al. (2016) Oil palm leaves and roots differ in physiological response, antioxidant enzyme activities and expression of stress-responsive genes upon exposure to drought stress. Acta Physiol Plantarum 38: 52. doi: 10.1007/s11738-016-2073-2
|
| [11] |
Silva PA, Oliveira IV, Rodrigues KCB, et al. (2016) Leaf gas exchange and multiple enzymatic and non-enzymatic antioxidant strategies related to drought tolerance in two oil palm hybrids. Trees 30: 203-214. doi: 10.1007/s00468-015-1289-x
|
| [12] | Suresh K, Nagamani C, Ramachandrudu K, et al. (2010) Gas-exchange characteristics, leaf water potential and chlorophyll a fluorescence in oil palm (Elaeis guineensis Jacq.) seedlings under water stress and recovery. Photosynthetica 48: 430-436. |
| [13] | Bahrami-Rad S, Hajiboland R (2017) Effect of potassium application in drought-stressed tobacco (Nicotiana rustica L.) plants: Comparison of root with foliar application. Ann Agric Sci 62: 121-130. |
| [14] |
Martineau E, Domec JC, Bosc A, et al. (2017) The effects of potassium nutrition on water use in field-grown maize (Zea mays L.). Environ Exp Botany 134: 62-71. doi: 10.1016/j.envexpbot.2016.11.004
|
| [15] |
Mohd Zain NA, Ismail MR (2016) Effects of potassium rates and types on growth, leaf gas exchange and biochemical changes in rice (Oryza sativa) planted under cyclic water stress. Agric Water Manage 164: 83-90. doi: 10.1016/j.agwat.2015.09.022
|
| [16] |
Zahoor R, Dong H, Abid M, et al. (2017) Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ Exp Botany 137: 73-83. doi: 10.1016/j.envexpbot.2017.02.002
|
| [17] |
Daliparthy J, Barker AV, Mondal SS (1994) Potassium fractions with other nutrients in crops: A review focusing on the tropics. J Plant Nutri 17: 1859-1886. doi: 10.1080/01904169409364852
|
| [18] | Hu W, Yang J, Meng Y, et al. (2015) Potassium application affects carbohydrate metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll and its relationship with boll biomass. Field Crops Res 179: 120-131. |
| [19] |
Hafeez A, Ali S, Ma X, et al. (2018) Potassium to nitrogen ratio favors photosynthesis in late-planted cotton at high planting density. Ind Crops Prod 124: 369-381. doi: 10.1016/j.indcrop.2018.08.006
|
| [20] | Mengel K (2016). Potassium. In Handbook of plant nutrition. CRC Press 107-136. |
| [21] |
Peoples TR, Koch DW (1979) Role of Potassium in Carbon Dioxide Assimilation in Medicago sativa L. Plant Physiol 63: 878-881. doi: 10.1104/pp.63.5.878
|
| [22] |
Sangakkara UR, Frehner M, Nösberger J (2000) Effect of soil moisture and potassium fertilizer on shoot water potential, photosynthesis and partitioning of carbon in mungbean and cowpea. J Agron Crop Sci 185: 201-207. doi: 10.1046/j.1439-037x.2000.00422.x
|
| [23] |
Ibrahim HM, Jaafar ZEH, Karimi E, et al. (2012) Primary, Secondary Metabolites, Photosynthetic Capacity and Antioxidant Activity of the Malaysian Herb Kacip Fatimah (Labisia Pumila Benth) Exposed to Potassium Fertilization under Greenhouse Conditions. Int J Mol Sci 13: 15321-15342. doi: 10.3390/ijms131115321
|
| [24] | Bocianowski J, Szulc P, Tratwal A, et al. (2016) The influence of potassium to mineral fertilizers on the maize health. J Int Agric15: 1286-1292. |
| [25] | Baque MA, Karim MA. Hamid A, et al. (2006) Effects of fertilizer potassium on growth, yield and nutrient uptake of wheat (Triticum aestivum) under water stress conditions. S Pacif Stud 27: 25-35. |
| [26] | Aboelill AA, Mehanna HM, Kassab OM, et al. (2012) The response of peanut crop to foliar spraying with potassium under water stress conditions. Aus J Basic Appl Sci 6: 626-634. |
| [27] | Vijiandran JR, Husni MHA. Teh CBS, et al. (2017) Nutrient losses through runoff from several types of fertilisers under mature oil palm. Malays J Soil Sci 21: 113-121. |
| [28] | Wahid MB, Simeh MA (2009) Issues related to production cost of palm oil in Malaysia. Oil Palm Indus Econ J 9: 1-12. |
| [29] | MPOB (2013) Oil Palm Biology: Facts & Figures. Kuala Lumpur: Malays Palm Oil Board 40-41. |
| [30] | Klapwijk D, Lint PJAL (1974) Fresh weight and flowering of tomato plants as influecend by container type and watering conditions. Acta Hortic 23: 237-248. |
| [31] | MPOB (2017) Oil palm Vegetative Measurements Manual. Kuala Lumpur. Malays Palm Oil Board 50-65. |
| [32] |
Weatherly P (1950) Studies in the water relations of the cotton plant. New Phytol 49: 81-97. doi: 10.1111/j.1469-8137.1950.tb05146.x
|
| [33] |
Jaafar HZE, Ibrahim MH, Fakri NFM (2012) Impact of soil field water capacity on secondary metabolites, phenylalanine ammonia-lyase (PAL), maliondialdehyde (MDA) and photosynthetic responses of Malaysian Kacip Fatimah (Labisia pumila Benth). Molecules 17: 7305-7322. doi: 10.3390/molecules17067305
|
| [34] | Zainuren NF, Ibrahim MH, Abdullah CAC, et al. (2019) Growth, Carbon Assimilation and Quality of Kesum (Persicaria minor) as Exposed to Zinc Oxide Nanoparticles. Annual Res Rev Biol 2019: 1-15. |
| [35] |
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant and Soil 39: 205-207. doi: 10.1007/BF00018060
|
| [36] | Ibrahim MH, Jaafar HZE, Rahmat A, et al. (2012) Involvement of Nitrogen on Flavonoids, Glutathione, Anthocyanin, Ascorbic Acid and Antioxidant Activities of Malaysian Medicinal Plant Labisia pumila Blume (Kacip Fatimah). Int J Mol Sci 13: 393-408. |
| [37] | Ibrahim MH, Jaafar HZE, Rahmat A, et al. (2011) The relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules 16: 162-174. |
| [38] |
Boutraa T, Akhkha A, Al-Shoaibi AA, et al. (2010) Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. J Taibah Univ Sci 3: 39-48. doi: 10.1016/S1658-3655(12)60019-3
|
| [39] |
Litvin AG, Van Iersel MW, Malladi A (2016) Drought stress reduces stem elongation and alters gibberellin-related gene expression during vegetative growth of tomato. J Am Soc Hortic Sci 141: 591-597. doi: 10.21273/JASHS03913-16
|
| [40] | Riaz A, Younis A, Taj AR, et al. (2013) Effect of drought stress on growth and flowering of marigold (Tagetes erecta L.). Pak J Bot 45: 123-131. |
| [41] |
Hussain M, Farooq S, Hasan W, et al. (2018) Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agri Water Manage 201: 152-166. doi: 10.1016/j.agwat.2018.01.028
|
| [42] | Anjum S, Xie X, Wang L, et al. (2011) Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agr Res 6: 2026-2032. |
| [43] | Rivera-Mendes YD, Cuenca JC, Romero HM (2016) Physiological responses of oil palm (Elaeis guineensis Jacq.) seedlings under different water soil conditions. Agrono Colombiana 34: 163. |
| [44] | Chartzoulakis K, Noitsakis B, Therios I (1993) Photosynthesis, plant growth and dry matter distribution in kiwifruit as influenced by water deficits. Irrigat Sci 14: 1-5. |
| [45] | Hailemichael G, Catalina A, González M, et al. (2016) Relationships between water status, leaf chlorophyll content and photosynthetic performance in Tempranillo vineyards. S Afr J Enol Vitic 37: 149-156. |
| [46] | Malika LY, Deshabandu KSHT, De Costa WAJM, et al. (2019) Physiological traits determining tolerance to intermittent drought in the Capsicum annuum complex. Sci Hortic246: 21-33. |
| [47] | Zhang S, Xu X, Sun Y, et al. (2018) Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J Int Agric17: 336-347. |
| [48] |
Oddo E, Inzerillo S, Grisafi F, et al. (2014) Does short-term potassium fertilization improve recovery from drought stress in laurel? Tree Physiol 34: 906-913. doi: 10.1093/treephys/tpt120
|
| [49] |
De Sá AFL, Valeri SV, Da Cruz MCP, et al. (2014) Effects of potassium application and soil moisture on the growth of Corymbia citriodora plants. Cerne 20: 645-651. doi: 10.1590/01047760201420041422
|
| [50] |
Fauzi WR, Putra ETS (2019) Dampak Pemberian Kalium Dan Cekaman Kekeringan Terhadap Serapan Hara Dan Produksi Biomassa Bibit Kelapa Sawit (Elaeis Gueenensis Jacq.). J Penelitian Kelapa Sawit 27: 41-56. doi: 10.22302/iopri.jur.jpks.v27i1.74
|
| [51] | Jordan-Meille L, Martineau E, Bornot Y, et al. (2018) How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions. Agriculture 8: 180. |
| [52] | Huda AI, El-Behairy UA, El-Desuki M, et al. (2010) Response of green bean to fertilization with potassium and magnesium. Res J Agric Biol Sci 6: 834-839. |
| [53] | Nazli F, Bushra, Iqbal MM, et al. (2018) Modeling the potassium requirements of potato crop for yield and quality optimization. Asian J Agric Biol 6: 169-180. |
| [54] |
Ortas I (2018) Influence of potassium and magnesium fertilizer application on the yield and nutrient accumulation of maize genotypes under field conditions. J Plant Nutri 41: 330-339. doi: 10.1080/01904167.2017.1385800
|
| [55] |
Chen H, Wang P, Li J, et al. (2012) Canopy Spectral Reflectance Feature and Leaf Water Potential of Sugarcane Inversion. Physics Procedia 25: 595-600. doi: 10.1016/j.phpro.2012.03.131
|
| [56] |
El-Hendawy SE, Al-Suhaibani NA, Elsayed S, et al. (2019) Potential of the existing and novel spectral reflectance indices for estimating the leaf water status and grain yield of spring wheat exposed to different irrigation rates. Agri Water Manage 217: 356-373. doi: 10.1016/j.agwat.2019.03.006
|
| [57] |
Ma X, He Q, Zhou G (2018) Sequence of Changes in Maize Responding to Soil Water Deficit and Related Critical Thresholds. Fron Plant Sci 9: 511. doi: 10.3389/fpls.2018.00511
|
| [58] |
Hsiao TC (1973) Plant responses to water stress. Ann Rev Plant Physio 24: 519-570. doi: 10.1146/annurev.pp.24.060173.002511
|
| [59] | Reddy PS (2019) "Breeding for Abiotic Stress Resistance in Sorghum." Breeding Sorghum for Diverse End Uses. Woodh Publ 2019: 325-340. |
| [60] | Ju Y, Yue X, Zhao X, et al. (2018) Physiological, micro-morphological and metabolomic analysis of grapevine (Vitis vinifera L.) leaf of plants under water stress. Plant Physiol Bioch 130: 501-510. |
| [61] | Jamaludin D, Abd Aziz S, Ahmad D, et al. (2015) Impedance analysis of Labisia pumila plant water status. Infor Process Agric 2: 161-168. |
| [62] |
Gautam A, Agrawal D, SaiPrasad SV, et al. (2014) A quick method to screen high and low yielding wheat cultivars exposed to high temperature. Physiol Mol Biol Plant 20: 533-537. doi: 10.1007/s12298-014-0252-4
|
| [63] |
Kalaji HM, Carpentier R, Allakhverdiev SI, et al. (2012) Fluorescence parameters as early indicators of light stress in barley. J Photoch Photobio 112: 1-6. doi: 10.1016/j.jphotobiol.2012.03.009
|
| [64] |
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence-a practical guide. J Exp Bot 51: 659-668. doi: 10.1093/jexbot/51.345.659
|
| [65] |
Živčák M, Brestič M, Olšovská K, et al. (2008) Performance index as a sensitive indicator of water stress in Triticum aestivum L. Plant Soil Environ 54: 133-139. doi: 10.17221/392-PSE
|
| [66] |
Liu B, Liang J, Tang G, et al. (2019) Drought stress affects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci Hortic 250: 230-235. doi: 10.1016/j.scienta.2019.02.056
|
| [67] |
Naidoo G, Naidoo KK (2018) Drought stress effects on gas exchange and water relations of the invasive weed Chromolaena odorata. Flora 248: 1-9. doi: 10.1016/j.flora.2018.08.008
|
| [68] |
Zain NAM, Ismail MR, Mahmood M, et al. (2014) Alleviation of water stress effects on mr220 rice by application of periodical water stress and potassium fertilization. Molecules 19: 1795-1819. doi: 10.3390/molecules19021795
|
| [69] |
Mohammadian R, Rahimian H, Moghaddam M, et al. (2003) The effect of early season drought on chlorophyll a fluorescence in sugar beet (Beta vulgaris L.). Pakistan J Biol Sci 6: 1763-1769. doi: 10.3923/pjbs.2003.1763.1769
|
| [70] |
Zlatev Z (2009) Drought-induced changes in chlorophyll fluorescence of young wheat plants. Biotech Biotec Equip 23: 438-441. doi: 10.1080/13102818.2009.10818458
|
| [71] |
Meng L, Song JF, Wen J, et al. (2016) Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 54: 414-421. doi: 10.1007/s11099-016-0191-0
|
| [72] |
Wang XM, Wang XK, Su YB, et al. (2019) Land pavement depresses photosynthesis in urban trees especially under drought stress. Sci Total Environ 653: 120-130. doi: 10.1016/j.scitotenv.2018.10.281
|
| [73] |
Li Y, Song H, Zhou L, et al. (2019) Tracking chlorophyll fluorescence as an indicator of drought and rewatering across the entire leaf lifespan in a maize field. Agri Water Manage 211: 190-201. doi: 10.1016/j.agwat.2018.09.050
|
| [74] |
Ma P, Bai T, Ma F (2015) Effects of progressive drought on photosynthesis and partitioning of absorbed light in apple trees. J Int Agric 14: 681-690. doi: 10.1016/S2095-3119(14)60871-6
|
| [75] | Gómez-del-Campo M, Ruiz C, Lissarrague JR (2002) Effect of water stress on leaf area development, photosynthesis, and productivity in Chardonnay and Airén grapevines. Am J Enol Viticult 53: 138-143. |
| [76] |
Zargar SM, Gupta N, Nazir M, et al. (2017) Impact of drought on photosynthesis: Molecular perspective. Plant Gene 11: 154-159. doi: 10.1016/j.plgene.2017.04.003
|
| [77] | Oosterhuis DM, Loka DA, Kawakam EM, et al. (2014) "The physiology of potassium in crop production." Advances in agronomy. Academic Press 126: 203-233. |
| [78] | Armstrong DL, Griffin KP (1998) Better crops with plant food. Int Pl Nutri Institute 3. |
| [79] | Vyas SP, Garg BK, Kathju S, et al. (2001) Influence of potassium on water relations, photosynthesis, nitrogen metabolism and yield of clusterbean under soil moisture stress. Indian J Plant Physiol 6: 30-37. |
| [80] | Farooq M, Hussain M, Wahid A, et al. (2012) Drought Stress in Plants: An Overview BT - Plant Responses to Drought Stress: From Morphological to Molecular Features. Berlin, Heidelberg. Springer Berlin Heidelberg 1-33. |
| [81] | Osakabe Y, Osakabe K, Shinozaki K, et al. (2014) Response of plants to water stress. Front Pl Sci 5: 86. |
| [82] |
Miyashita K, Tanakamaru S, Maitani T, et al. (2005) Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ Exp Botany 53: 205-214. doi: 10.1016/j.envexpbot.2004.03.015
|
| [83] | Matthews LR (1986) The effect of water stress on the leaf area of Cucumis melo (Master thesis).Texas Tech University. Lubbock. United states. |
| [84] | Klamkowski K, Treder W (2008) Response to drought stress of three strawberry cultivars grown under greenhouse conditions. J Fruit Ornam Pl Res 16: 179-188. |
| [85] | Bolat I, Dikilitas M, Ercisli S, et al. (2014) The effect of water stress on some morphological, physiological, and biochemical characteristics and bud success on apple and quince rootstocks. The Sci World J 2014: 769732. |
| [86] |
Tominaga J, Shimada H, Kawamitsu Y (2018) Direct measurement of intercellular CO2 concentration in a gas-exchange system resolves overestimation using the standard method. J Exp Bot 69: 1981-1991. doi: 10.1093/jxb/ery044
|
| [87] |
Kelly JWG, Duursma RA, Atwell BJ, et al. (2016) Drought×CO2 interactions in trees: a test of the low-intercellular CO2 concentration (Ci) mechanism. New Phytol 209: 1600-1612. doi: 10.1111/nph.13715
|
| [88] |
Warren CR, Livingston NJ, Turpin DH (2004) Water stress decreases the transfer conductance of Douglas-fir (Pseudotsuga menziesii) seedlings. Tree Physiol 24: 971-979. doi: 10.1093/treephys/24.9.971
|
| [89] |
Guo YY, Yu HY, Yang MM, et al. (2018) Effect of Drought Stress on Lipid Peroxidation, Osmotic Adjustment and Antioxidant Enzyme Activity of Leaves and Roots of Lycium ruthenicum Murr. Seedling. Russ J Plant Physiol 65: 244-250. doi: 10.1134/S1021443718020127
|
| [90] | Toscano S, Farieri E, Ferrante A, et al. (2016) Physiological and Biochemical Responses in Two Ornamental Shrubs to Drought Stress. Front Plant Sci 7: 645. |
| [91] | Chrysargyris A, Drouza C, Tzortzakis N (2017) Optimization of potassium fertilization/nutrition for growth, physiological development, essential oil composition and antioxidant activity of Lavandula angustifolia Mill. J Soil Sci Plant Nutri 17: 291-306. |
| [92] | Siddiqui MH, Al-Whaibi MH, Sakran AM, et al. (2012) Effect of calcium and potassium on antioxidant system of Vicia faba L. Under cadmium stress. Int J Mol Sci 13: 6604-6619. |
| [93] | Su D, Chen N, Gao T, et al. (2012) Effects of Si+, K+, and Ca2+ on antioxidant enzyme activities and osmolytes in Halocnemum strobilaceum under salt stress. Adv Mat Res 56: 2542-2550. |
| [94] | Hasanuzzaman M, Bhuyan MHMB, Nahar K, et al. (2018) Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agron 8. |
| [95] |
Ma L, Shi Y (2011) Effects of potassium fertilizer on physiological and biochemical index of Stevia rebaudiana Bertoni. Energ Procedia 5: 581-586. doi: 10.1016/j.egypro.2011.03.102
|
| [96] | El-Mogy MM, Salama AM, Mohamed HFY, et al. (2019) Responding of long green pepper plants to different sources of foliar potassium fertiliser. Agriculture 65: 59-76. |
| [97] |
Nguyen PM, Kwee EM, Niemeyer ED (2010) Potassium rate alters the antioxidant capacity and phenolic concentration of basil (Ocimum basilicum L.) leaves. Food Chem 123: 1235-1241. doi: 10.1016/j.foodchem.2010.05.092
|
| [98] | Zhang W, Zhang X, Wang Y, et al. (2018) Potassium fertilization arrests malate accumulation and alters soluble sugar metabolism in apple fruit. Biol Open 7. |
| [99] |
Klunklin W, Savage G (2017) Effect on Quality Characteristics of Tomatoes Grown Under Well-Watered and Drought Stress Conditions. Foods 6: 56. doi: 10.3390/foods6080056
|
| [100] |
Mathesius U (2018) Flavonoid Functions in Plants and Their Interactions with Other Organisms. Plants 7: 30. doi: 10.3390/plants7020030
|
| [101] | Yuan Y, Liu Y, Wu C, et al. (2012) Water Deficit Affected Flavonoid Accumulation by Regulating Hormone Metabolism in Scutellaria baicalensis Georgi Roots. Plos One 7. |
| [102] |
Munene R, Changamu E, Korir N, et al. (2017) Effects of different nitrogen forms on growth, phenolics, flavonoids and antioxidant activity in amaranth species. Trop Plant Res 4: 81-89. doi: 10.22271/tpr.2017.v4.i1.012
|
| [103] |
Salas-Pérez L, Fornari-Reale T, Preciado-Rangel P, et al. (2018) Cultivar Variety and Added Potassium Influence the Nutraceutical and Antioxidant Content in Hydroponically Grown Basil (Ocimum basilicum L.). Agron 8: 13. doi: 10.3390/agronomy8020013
|
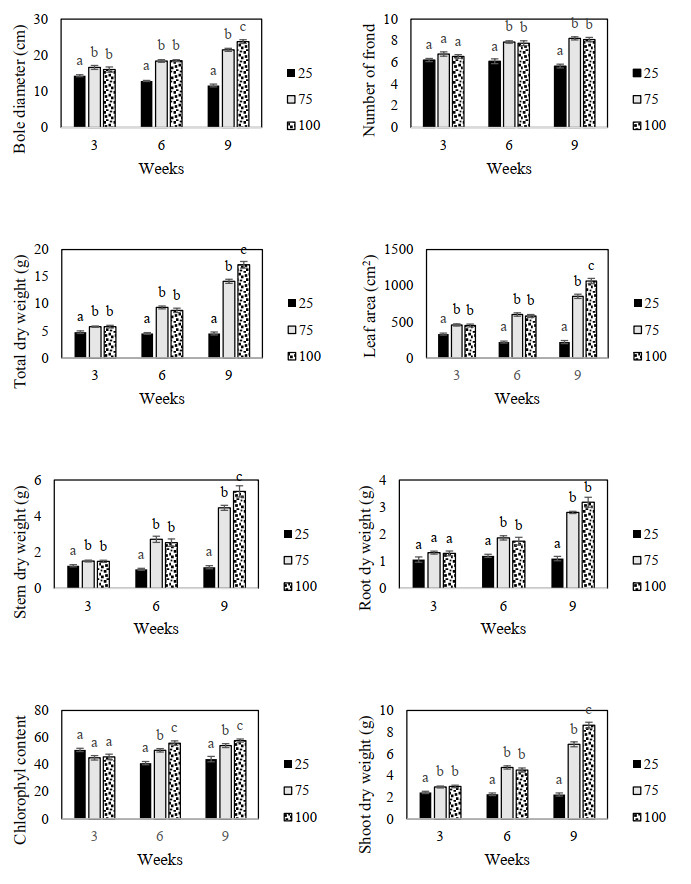
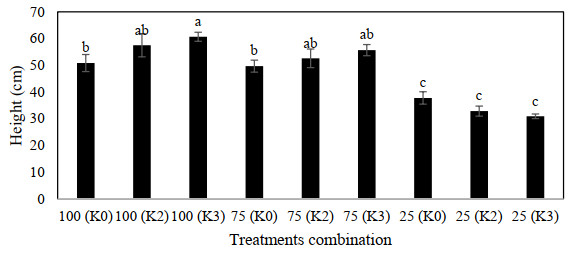
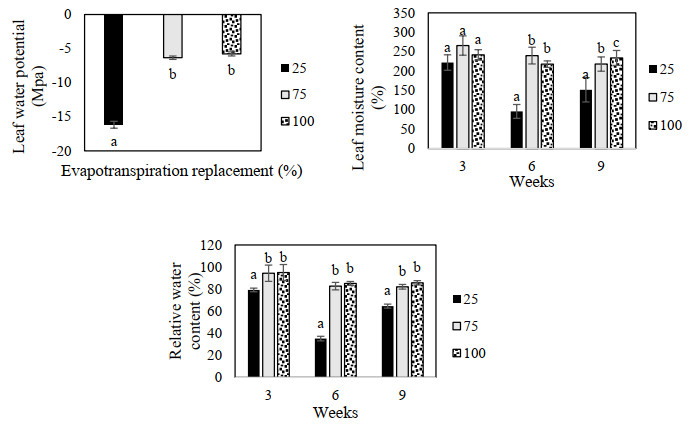
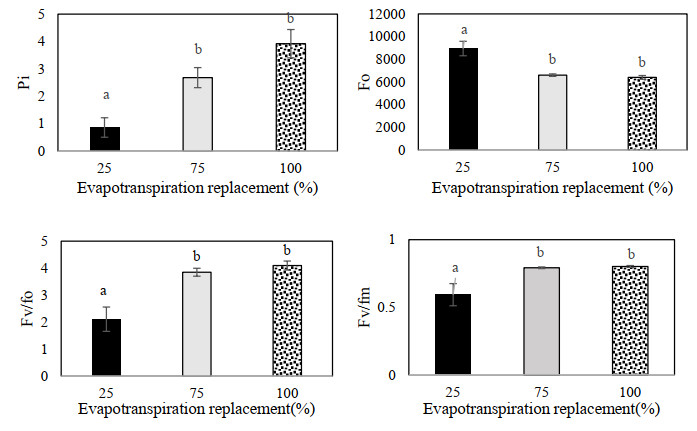
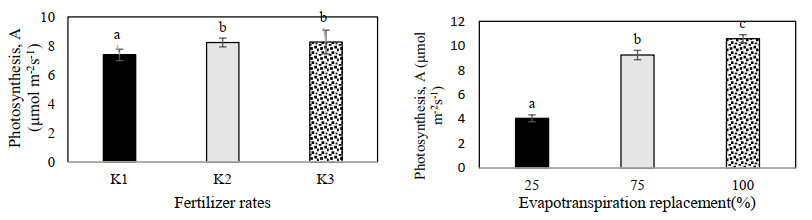
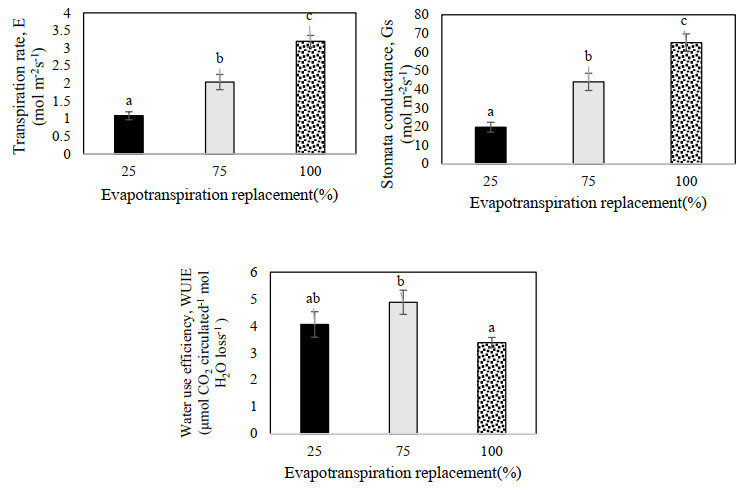
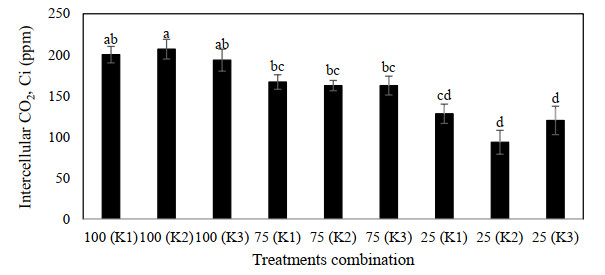
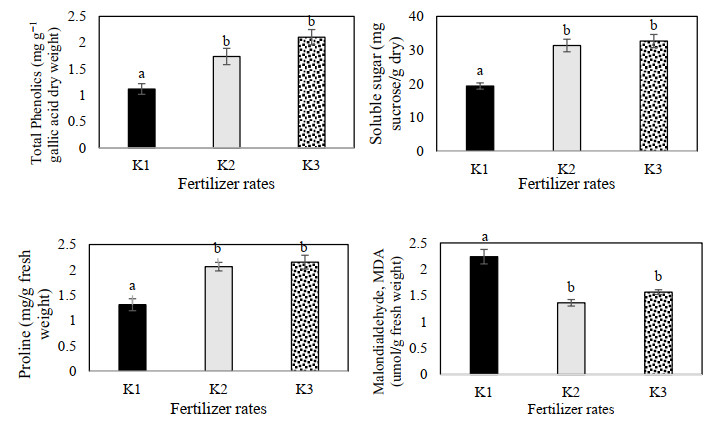
Figures(9) / Tables(3)

Tuan Syaripah Najihah, Mohd Hafiz Ibrahim, Nurul Amalina Mohd Zain, Rosimah Nulit, Puteri Edaroyati Megat Wahab. Activity of the oil palm seedlings exposed to a different rate of potassium fertilizer under water stress condition[J]. AIMS Environmental Science, 2020, 7(1): 46-68. doi: 10.3934/environsci.2020004







 DownLoad:
DownLoad: