Citation: Siming Liu, Honglei Gao, Peng Hou, Yong Tan. Risk spillover effects of international crude oil market on China’s major markets[J]. AIMS Energy, 2019, 7(6): 819-840. doi: 10.3934/energy.2019.6.819

| [1] |
Balcilar M, Hammoudeh S, Toparli E (2018) On the risk spillover across the oil market, stock market, and the oil related CDS sectors: A volatility impulse response approach. Energy Econ 74: 813-827. doi: 10.1016/j.eneco.2018.07.027

|
| [2] |
Bouri E (2015) A broadened causality in variance approach to assess the risk dynamics between crude oil prices and the jordanian stock market. Energy Policy 85: 271-279. doi: 10.1016/j.enpol.2015.06.001
|
| [3] |
Ji Q, Liu B, Fan Y (2019) Risk dependence of CoVaR and structural change between oil prices and exchange rates: A time-varying copula model. Energy Econ 77: 80-92. doi: 10.1016/j.eneco.2018.07.012
|
| [4] |
Li X, Wei Y (2018) The dependence and risk spillover between crude oil market and China stock market: New evidence from a variational mode decomposition-based copula method. Energy Econ 74: 565-581. doi: 10.1016/j.eneco.2018.07.011
|
| [5] |
Mensi W, Hammoudeh S, Shahzad S, et al. (2017) Oil and foreign exchange market tail dependence and risk spillovers for MENA, emerging and developed countries: VMD decomposition based copulas. Energy Econ 67: 476-495. doi: 10.1016/j.eneco.2017.08.036
|
| [6] |
Mensi W, Hammoudeh S, Shahzad S, et al. (2017) Modeling systemic risk and dependence structure between oil and stock markets using a variational mode decomposition-based copula method. J Bank Financ 75: 258-279. doi: 10.1016/j.jbankfin.2016.11.017
|
| [7] |
Pavlova I, De Boyrie ME, Parhizgari AM (2018) A dynamic spillover analysis of crude oil effects on the sovereign credit risk of exporting countries. Q Rev Econ Financ 68: 10-22. doi: 10.1016/j.qref.2018.03.003
|
| [8] |
Shahzad S, Mensi W, Hammoudeh S, et al. (2018) Extreme dependence and risk spillovers between oil and Islamic stock markets. Emerg Mark Rev 34: 42-63. doi: 10.1016/j.ememar.2017.10.003
|
| [9] |
Wen D, Wang GJ, Ma C, et al. (2019) Risk spillovers between oil and stock markets: A VAR for VaR analysis. Energy Econ 80: 524-535. doi: 10.1016/j.eneco.2019.02.005
|
| [10] |
Du L, He Y (2015) Extreme risk spillovers between crude oil and stock markets. Energy Econ 51: 455-465. doi: 10.1016/j.eneco.2015.08.007
|
| [11] |
Hong Y, Liu Y, Wang S (2009) Granger causality in risk and detection of extreme risk spillover between financial markets. J Econometrics 150: 271-287. doi: 10.1016/j.jeconom.2008.12.013
|
| [12] |
Elyasiani E, Gambarelli L, Muzzioli S (2017) The information content of corridor volatility measures during calm and turmoil periods. Quant Financ Econ 1: 454-473. doi: 10.3934/QFE.2017.4.454
|
| [13] |
Hamao Y, Masulis RW, Ng V (1990) Correlations in price changes and volatility across international stock markets. Rev Financ Stud 3: 281-307. doi: 10.1093/rfs/3.2.281
|
| [14] |
Hong Y (2001) A test for volatility spillover with applications to exchange rates. J Econometrics 103: 183-224. doi: 10.1016/S0304-4076(01)00043-4
|
| [15] |
Madaleno M, Vieira E (2018) Volatility analysis of returns and risk: family versus nonfamily firms. Quant Financ Econ 2: 348-372. doi: 10.3934/QFE.2018.2.348
|
| [16] |
Kang W, Ratti RA, Yoon KH (2015) The impact of oil price shocks on the stock market return and volatility relationship. J Int Financ Mark Inst Money 34: 41-54. doi: 10.1016/j.intfin.2014.11.002
|
| [17] |
Xu W, Ma F, Chen W, et al. (2019) Asymmetric volatility spillovers between oil and stock markets: Evidence from China and the United States. Energy Econ 80: 310-320. doi: 10.1016/j.eneco.2019.01.014
|
| [18] |
Peng C, Zhu H, Guo Y, et al. (2018) Risk spillover of international crude oil to China's firms: Evidence from granger causality across quantile. Energy Econ 72: 188-199. doi: 10.1016/j.eneco.2018.04.007
|
| [19] |
Zhang J (2019) Oil and gas trade between China and countries and regions along the 'Belt and Road': A panoramic perspective. Energy Policy 129: 1111-1120. doi: 10.1016/j.enpol.2019.03.020
|
| [20] |
Ren YH, Gu WW (2017) The impact on RMB internationalization process: Exchange rate expectation and foreign trade. Quant Financ Econ 1:114-124. doi: 10.3934/QFE.2017.1.114
|
| [21] | Babalos V, Stavroyiannis S, Gupta R (2015) Do commodity investors herd? Evidence from a time-varying stochastic volatility model. Resour Policy 46: 281-287. |
| [22] |
Irwin SH, Sanders DR (2011) Index funds, financialization, and commodity futures markets. Appl Econ Perspect Policy 33: 1-31. doi: 10.1093/aepp/ppq032
|
| [23] |
Tang K, Xiong W (2012) Index investment and the financialization of commodities. Financ Anal J 68: 54-74. doi: 10.2469/faj.v68.n6.5
|
| [24] |
Wen S, An H, Huang S, et al. (2019) Dynamic impact of China's stock market on the international commodity market. Resour Policy 61: 564-571. doi: 10.1016/j.resourpol.2018.06.009
|
| [25] | Zhang YJ, Chevallier J, Guesmi K (2017) 'De‐financialization' of commodities? Evidence from stock, crude oil and natural gas markets. Energy Econ 68: 228-239. |
| [26] |
Adams Z, Glück T (2015) Financialization in commodity markets: A passing trend or the new normal? J Bank Financ 60: 93-111. doi: 10.1016/j.jbankfin.2015.07.008
|
| [27] | Cheng IH, Xiong W (2013) The financialization of commodity markets. NBER Working Paper No.19642. |
| [28] |
Diebold FX, Yilmaz K (2012) Better to give than to receive: Predictive directional measurement of volatility spillovers. Int J Forecasting 28: 57-66. doi: 10.1016/j.ijforecast.2011.02.006
|
| [29] |
Hamilton JD (1983) Oil and the macroeconomy since World War II. J Polit Econ 91: 228-248. doi: 10.1086/261140
|
| [30] |
An Y, Sun M, Gao C, et al. (2018) Analysis of the impact of crude oil price fluctuations on China's stock market in different periods-Based on time series network model. Physica A 492: 1016-1031. doi: 10.1016/j.physa.2017.11.032
|
| [31] |
Atems B, Kapper D, Lam E (2015) Do exchange rates respond asymmetrically to shocks in the crude oil market? Energy Econ 49: 227-238. doi: 10.1016/j.eneco.2015.01.027
|
| [32] |
Bildirici ME, Badu MM (2018) The effects of oil prices on confidence and stock return in China, India and Russia. Quant Financ Econ 2: 884-903. doi: 10.3934/QFE.2018.4.884
|
| [33] | Chen H, Liu L, Wang Y, et al. (2016) Oil price shocks and U.S. dollar exchange rates. Energy 112: 1036-1048. |
| [34] |
Ghosh S, Kanjilal K (2016) Co-movement of international crude oil price and Indian stock market: evidences from nonlinear cointegration tests. Energy Econ 53: 111-117. doi: 10.1016/j.eneco.2014.11.002
|
| [35] |
Kang SH, Mciver R, Yoon SM (2017) Dynamic spillover effects among crude oil, precious metal, and agricultural commodity futures markets. Energy Econ 62: 19-32. doi: 10.1016/j.eneco.2016.12.011
|
| [36] |
Khalfaoui R, Boutahar M, Boubaker H (2015) Analyzing volatility spillovers and hedging between oil and stock markets: Evidence from wavelet analysis. Energy Econ 49: 540-549. doi: 10.1016/j.eneco.2015.03.023
|
| [37] |
Luo J, Ji Q (2018) High-frequency volatility connectedness between the US crude oil market and China's agricultural commodity markets. Energy Econ 76: 424-438. doi: 10.1016/j.eneco.2018.10.031
|
| [38] |
Mensi W, Beljid M, Boubaker A, et al. (2013) Correlations and volatility spillovers across commodity and stock markets: Linking energies, food, and gold. Econ Model 32: 15-22. doi: 10.1016/j.econmod.2013.01.023
|
| [39] |
Turhan MI, Sensoy A, Hacihasanoglu E (2014) A comparative analysis of the dynamic relationship between oil prices and exchange rates. J Int Financ Mark Inst Money 32: 397-414. doi: 10.1016/j.intfin.2014.07.003
|
| [40] |
Ji Q, Bouri E, Roubaud D, et al. (2018) Risk spillover between energy and agricultural commodity markets: A dependence-switching CoVaR-copula model. Energy Econ 75: 14-27. doi: 10.1016/j.eneco.2018.08.015
|
| [41] |
Algieri B, Leccadito A (2017) Assessing contagion risk from energy and non-energy commodity markets. Energy Econ 62: 312-322. doi: 10.1016/j.eneco.2017.01.006
|
| [42] |
Wang GJ, Xie C, Jiang ZQ, et al. (2016) Who are the net senders and recipients of volatility spillovers in China's financial markets? Financ Res Lett 18: 255-262. doi: 10.1016/j.frl.2016.04.025
|
| [43] |
Demirer M, Diebold FX, Liu L, et al. (2018). Estimating global bank network connectedness. J App Econom 33: 1-15. doi: 10.1002/jae.2585
|
| [44] |
Wang GJ, Xie C, Zhao LF, et al. (2018) Volatility connectedness in the Chinese banking system: Do state-owned commercial banks contribute more? J In Financ Mark Inst Money 57: 205-230. doi: 10.1016/j.intfin.2018.07.008
|
| [45] |
Yi SY, Xu ZS, Wang GJ (2018) Volatility connectedness in the cryptocurrency market: Is Bitcoin a dominant cryptocurrency? Int Rev Financ Anal 60: 98-114. doi: 10.1016/j.irfa.2018.08.012
|
| [46] |
Koop G, Pesaran MH, Potter SM (1996) Impulse response analysis in nonlinear multivariate models. J Econometrics 74:119-147. doi: 10.1016/0304-4076(95)01753-4
|
| [47] |
Pesaran MH, Shin Y (1998) Generalised impulse response analysis in linear multivariate models. Econ Lett 58: 17-29. doi: 10.1016/S0165-1765(97)00214-0
|
| [48] |
Engle RF, Manganelli S (2004) CAViaR: Conditional autoregressive value at risk by regression quantiles. J Bus Econ Stat 22: 367-381. doi: 10.1198/073500104000000370
|
| [49] | Qureshi K (2016) Value-at-Risk: The effect of autoregression in a quantile process. arXiv preprint arXiv: 1605.04940. |
| [50] |
Bernardi M, Catania L (2016) Comparison of Value-at-Risk models using the MCS approach. Comput Stat 31: 579-608. doi: 10.1007/s00180-016-0646-6
|
| [51] |
Ferraty F, Quintela-Del-Río A (2016) Conditional VAR and expected shortfall: A new functional approach. Econ Rev 35: 263-292. doi: 10.1080/07474938.2013.807107
|
| [52] |
Gerlach R, Lu ZD, Huang H (2013) Exponentially smoothing the skewed laplace distribution for Value-at-Risk forecasting. J Forecasting 32: 534-550. doi: 10.1002/for.2255
|
| [53] |
Gkillas K, Katsiampa P (2018) An application of extreme value theory to cryptocurrencies. Econ Lett 164: 109-111. doi: 10.1016/j.econlet.2018.01.020
|
| [54] |
Li ZH, Dong H, Huang ZH, et al. (2018) Asymmetric effects on risks of Virtual Financial Assets (VFAs) in different regimes: A case of bitcoin. Quant Financ Econ 2: 860-883. doi: 10.3934/QFE.2018.4.860
|
| [55] |
Righi MB, Ceretta PS (2015) Forecasting value at risk and expected shortfall based on serial pair-copula constructions. Expert Sys Appl 42: 6380-6390. doi: 10.1016/j.eswa.2015.04.023
|
| [56] |
Whitea H, Kimb TH, Manganelli S (2015) VAR for VaR: Measuring tail dependence using multivariate regression quantiles. J Econometrics 187: 169-188. doi: 10.1016/j.jeconom.2015.02.004
|
| [57] |
Gong X, Wen F, Xia XH, et al. (2017) Investigating the risk-return trade-off for crude oil futures using high-frequency data. Appl Energy 196: 152-161. doi: 10.1016/j.apenergy.2016.11.112
|
| [58] | Chen SL, Liu SM, Cai RJ, et al. (2019) The factors that influence exchange-rate risk: Evidence in China, Emerg Mark Financ Tr. |
| [59] |
Dong H, Liu Y, Chang JQ (2019) The heterogeneous linkage of economic policy uncertainty and oil return risks. Green Financ 1: 46-66. doi: 10.3934/GF.2019.1.46
|
| [60] |
Liao GK, Li ZH, Du ZQ, et al. (2019) The heterogeneous interconnections between supply or demand side and oil risks. Energies 12: 2226. doi: 10.3390/en12112226
|
| [61] |
An Y, Sun M, Gao CX, et al. (2018) Analysis of the impact of crude oil price fluctuations on China's stock market in different periods-based on time series network model. Physica A 492: 1016-1031. doi: 10.1016/j.physa.2017.11.032
|
| [62] |
Arouri MEH, Jouini J, Nguyen DK (2012) On the impacts of oil price fluctuations on European equity markets: Volatility spillover and hedging effectiveness. Energy Econ 34: 611-617. doi: 10.1016/j.eneco.2011.08.009
|
| [63] |
Zhang YL, Wang J (2019) Linkage influence of energy market on financial market by multiscale complexity synchronization. Physica A 516: 254-266. doi: 10.1016/j.physa.2018.10.038
|
| [64] |
Guo J (2018) Co-movement of international copper prices, China's economic activity, and stock returns: Structural breaks and volatility dynamics. Glob Financ J 36: 62-77. doi: 10.1016/j.gfj.2018.01.001
|
| [65] |
Zheng X, Chen BM (2014) Identification of stock market forces in the system adaptation framework. Inform Sci 265: 105-122. doi: 10.1016/j.ins.2013.12.028
|
| [66] |
Zhang C, Chen X (2014) The impact of global oil price shocks on China's bulk commodity markets and fundamental industries. Energy Policy 66: 32-41. doi: 10.1016/j.enpol.2013.09.067
|
| [67] |
Burdekin RCK, Siklos PL (2018) Quantifying the impact of the November 2014 Shanghai-Hong Kong stock connect. Int Rev Econ Financ 57: 156-163. doi: 10.1016/j.iref.2018.01.001
|
| [68] |
Ruan Q, Bao J, Zhang M, et al. (2019) The effects of exchange rate regime reform on RMB markets: A new perspective based on MF-DCCA. Physica A 522: 122-134. doi: 10.1016/j.physa.2019.01.110
|
| [69] |
Ahmad W, Rais S, Shaik AR (2018) Modelling the directional spillovers from DJIM Index to conventional benchmarks: Different this time? Q Rev Econ Financ 67: 14-27. doi: 10.1016/j.qref.2017.04.012
|
| [70] |
Antonakakis N, Kizys R (2015) Dynamic spillovers between commodity and currency markets. Int Rev Financ Anal 41: 303-319. doi: 10.1016/j.irfa.2015.01.016
|
| [71] |
Gillaizeau M, Jayasekera R, Maaitah A, et al. (2019) Giver and the receiver: Understanding spillover effects and predictive power in cross-market Bitcoin prices. Int Rev Financ Anal 63: 86-104. doi: 10.1016/j.irfa.2019.03.005
|
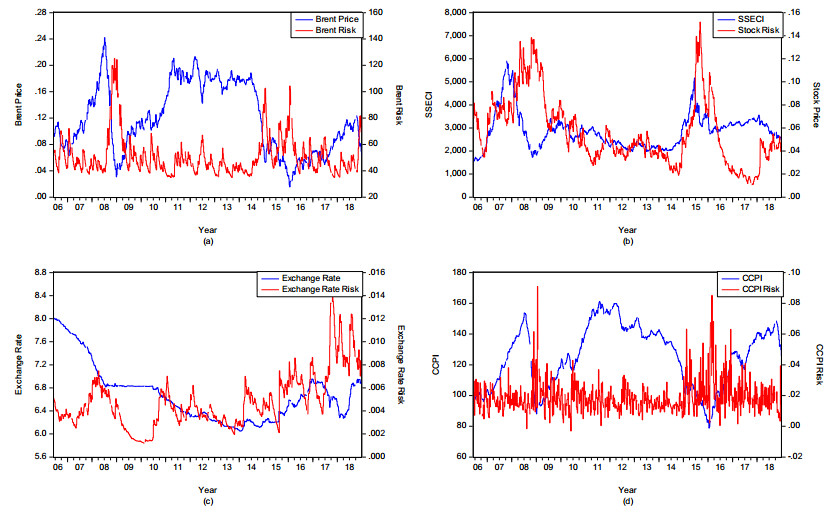
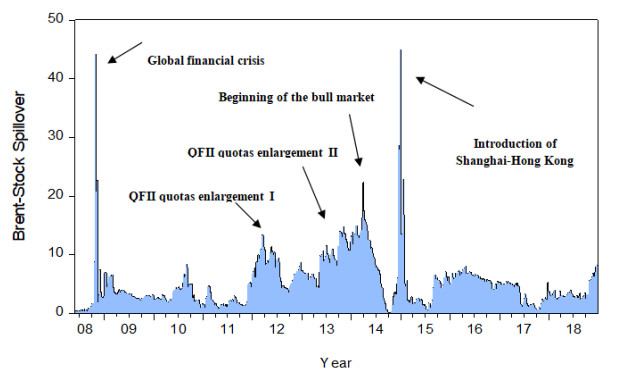
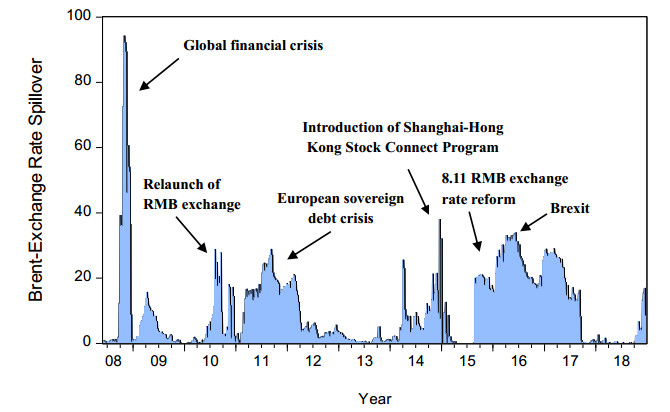
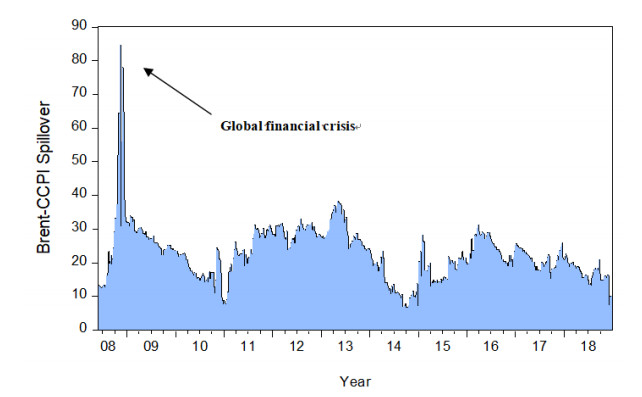
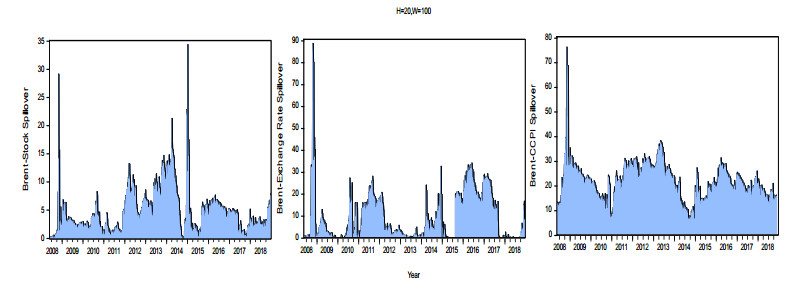
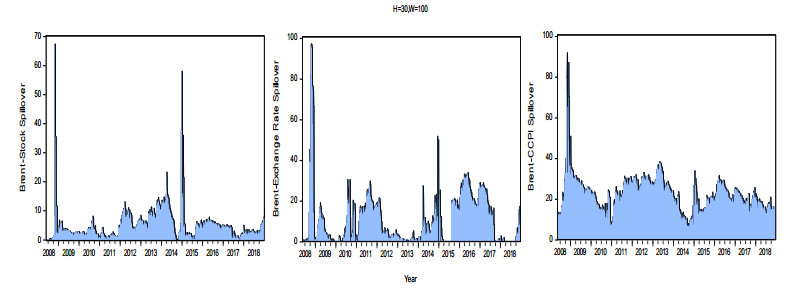
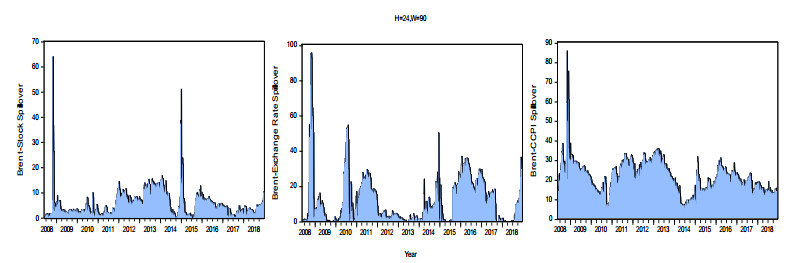
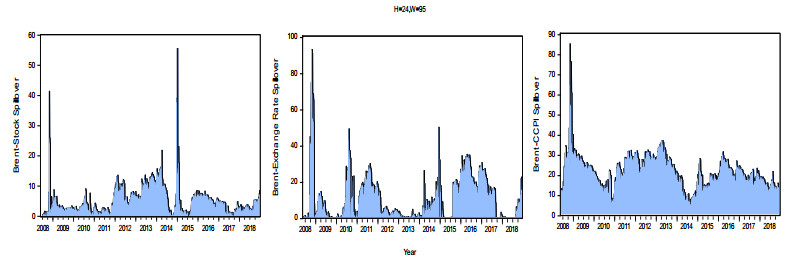
Figures(8) / Tables(3)

Siming Liu, Honglei Gao, Peng Hou, Yong Tan. Risk spillover effects of international crude oil market on China’s major markets[J]. AIMS Energy, 2019, 7(6): 819-840. doi: 10.3934/energy.2019.6.819







 DownLoad:
DownLoad: