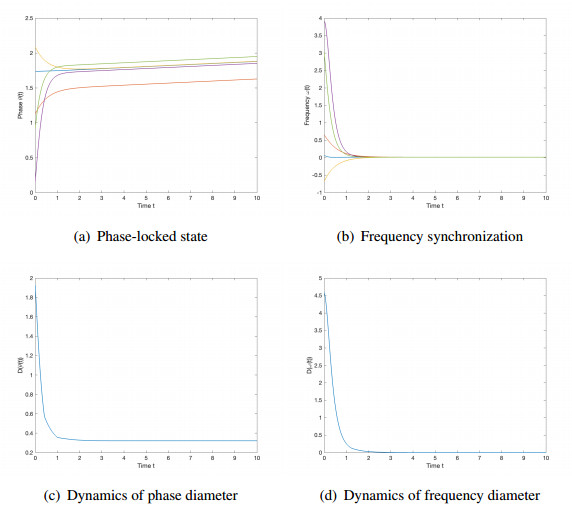
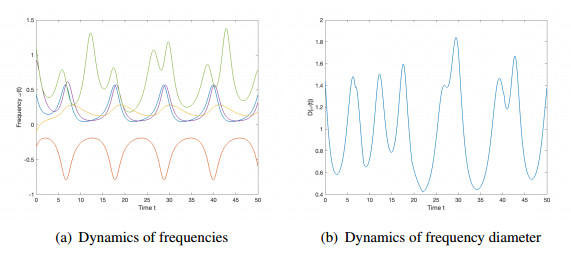
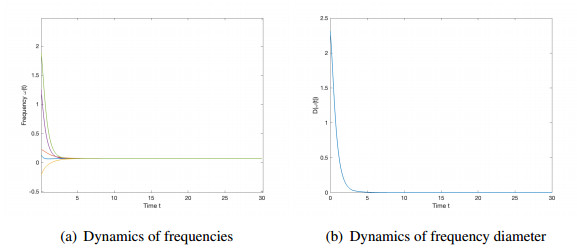
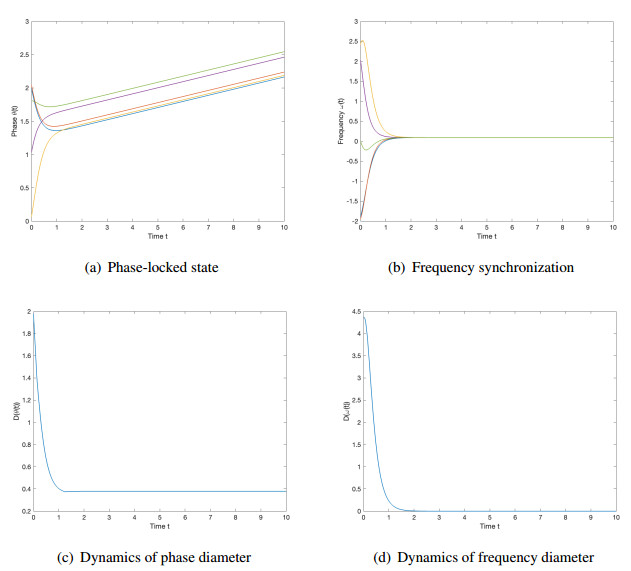
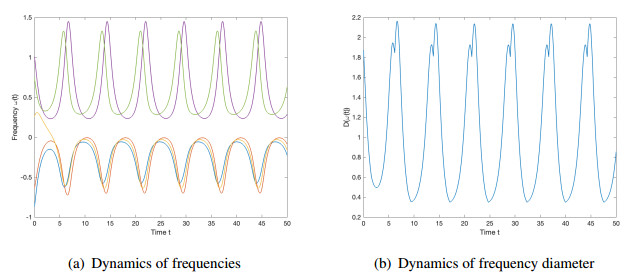
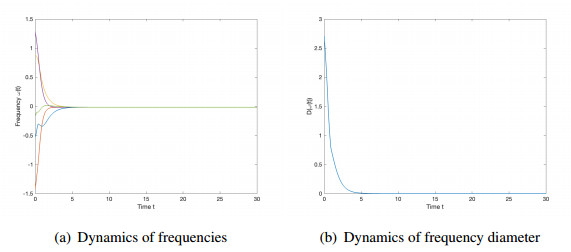
We studied the collective behaviors of the time-delayed Kuramoto model with frustration under general network topology. For the generalized Kuramoto model with the graph diameter no greater than two and under a sufficient regime in terms of small time delay and frustration and large coupling strength, we showed that the complete frequency synchronization occurs exponentially fast when the initial configuration is distributed in a half circle. We also studied a complete network, which is a small perturbation of all-to-all coupling, as well as presented sufficient frameworks leading to the exponential emergence of frequency synchronization for the initial data confined in a half circle.
Citation: Tingting Zhu. Synchronization of the generalized Kuramoto model with time delay and frustration[J]. Networks and Heterogeneous Media, 2023, 18(4): 1772-1798. doi: 10.3934/nhm.2023077
We studied the collective behaviors of the time-delayed Kuramoto model with frustration under general network topology. For the generalized Kuramoto model with the graph diameter no greater than two and under a sufficient regime in terms of small time delay and frustration and large coupling strength, we showed that the complete frequency synchronization occurs exponentially fast when the initial configuration is distributed in a half circle. We also studied a complete network, which is a small perturbation of all-to-all coupling, as well as presented sufficient frameworks leading to the exponential emergence of frequency synchronization for the initial data confined in a half circle.

| [1] |
J. A. Acebrón, L. L. Bonilla, C. J. P. Vicente, F. Ritort, R. Spigler, The Kuramoto model: A simple paradigm for synchronization phenomena, Rev. Mod. Phys., 77 (2005), 137–185. https://doi.org/10.1103/RevModPhys.77.137 doi: 10.1103/RevModPhys.77.137

|
| [2] |
J. Buck, E. Buck, Biology of synchronous flashing of fireflies, Nature, 211 (1966), 562–564. https://doi.org/10.1038/211562a0 doi: 10.1038/211562a0
|
| [3] | F. Bullo, J. Cortés, S. Martínez, Distributed Control of Robotic Networks: A Mathematical Approach to Motion Coordination Algorithms, Princeton: Princeton University Press, 2009. |
| [4] | S. Camazine, J. L. Deneubourg, N. R. Franks, J. Sneyd, G. Theraulaz, E. Bonabeau, Self-Organization in Biological Systems, Princeton: Princeton University Press, 2001. |
| [5] |
Y. P. Choi, S. Y. Ha, S. Jung, Y. Kim, Asymptotic formation and orbital stability of phase-locked states for the Kuramoto model, Phys. D, 241 (2012), 735–754. https://doi.org/10.1016/j.physd.2011.11.011 doi: 10.1016/j.physd.2011.11.011
|
| [6] |
M. Y. Choi, H. J. Kim, D. Kim, Synchronization in a system of globally coupled oscillators with time delay, Phys. Rev. E, 61 (2000), 371–381. https://doi.org/10.1103/PhysRevE.61.371 doi: 10.1103/PhysRevE.61.371
|
| [7] |
Y. P. Choi, C. Pignotti, Exponential synchronization of Kuramoto oscillators with time delayed coupling, Commun. Math. Sci., 19 (2021), 1429–1445. https://dx.doi.org/10.4310/CMS.2021.v19.n5.a11 doi: 10.4310/CMS.2021.v19.n5.a11
|
| [8] |
N. Chopra, M. W. Spong, On exponential synchronization of Kuramoto oscillators, IEEE Trans. Automat. Control, 54 (2009), 353–357. https://doi.org/10.1109/TAC.2008.2007884 doi: 10.1109/TAC.2008.2007884
|
| [9] |
H. Daido, Quasientrainment and slow relaxation in a population of oscillators with random and frustrated interactions, Phys. Rev. Lett., 68 (1992), 1073–1076. https://doi.org/10.1103/PhysRevLett.68.1073 doi: 10.1103/PhysRevLett.68.1073
|
| [10] |
J. G. Dong, S. Y. Ha, D. Kim, Emergent Behavior of the Kuramoto model with a time delay on a general digraph, SIAM J. Appl. Dyn. Syst., 19 (2020), 304–328. https://doi.org/10.1137/19M1249096 doi: 10.1137/19M1249096
|
| [11] |
J. G. Dong, X. Xue, Synchronization analysis of Kuramoto oscillators, Commun. Math. Sci., 11 (2013), 465–480. https://dx.doi.org/10.4310/CMS.2013.v11.n2.a7 doi: 10.4310/CMS.2013.v11.n2.a7
|
| [12] |
S. Y. Ha, Y. Kim, Z. Li, Large-time dynamics of Kuramoto oscillators under the effects of inertia and frustration, SIAM J. Appl. Dyn. Syst., 13 (2014), 466–492. https://doi.org/10.1137/130926559 doi: 10.1137/130926559
|
| [13] |
S. Y. Ha, H. K. Kim, J. Park, Remarks on the complete synchronization of Kuramoto oscillators, Nonlinearity, 28 (2015), 1441–1462. http://dx.doi.org/10.1088/0951-7715/28/5/1441 doi: 10.1088/0951-7715/28/5/1441
|
| [14] |
S. Y. Ha, H. K. Kim, J. Park, Remarks on the complete synchronization for the Kuramoto model with frustrations, Anal. Appl., 16 (2018), 525–563. https://doi.org/10.1142/S0219530517500130 doi: 10.1142/S0219530517500130
|
| [15] |
S. Y. Ha, H. K. Kim, S. W. Ryoo, Emergence of phase-locked states for the Kuramoto model in a large coupling regime, Commun. Math. Sci., 14 (2016), 1073–1091. https://dx.doi.org/10.4310/CMS.2016.v14.n4.a10 doi: 10.4310/CMS.2016.v14.n4.a10
|
| [16] |
S. Y. Ha, D. Ko, Y. Zhang, Emergence of phase-locking in the Kuramoto model for identical oscillators with frustration, SIAM J. Appl. Dyn. Syst., 17 (2018), 581–625. https://doi.org/10.1137/17M1112959 doi: 10.1137/17M1112959
|
| [17] |
S. Y. Ha, Z. Li, Complete synchronization of Kuramoto oscillators with hierarchical leadership, Commun. Math. Sci., 12 (2014), 485–508. https://dx.doi.org/10.4310/CMS.2014.v12.n3.a5 doi: 10.4310/CMS.2014.v12.n3.a5
|
| [18] |
S. Y. Ha, Z. Li, X. Xue, Formation of phase-locked states in a population of locally interacting Kuramoto oscillators, J. Differ. Equ., 255 (2013), 3053–3070. https://doi.org/10.1016/j.jde.2013.07.013 doi: 10.1016/j.jde.2013.07.013
|
| [19] | J. K. Hale, S. M. V. Lunel, Introduction to functional-differential equations, in Applied Mathematical Sciences, New York: Springer-Verlag, 99 (1993). https://doi.org/10.1007/978-1-4612-4342-7 |
| [20] |
C. H. Hsia, C. Y. Jung, B. Kwon, Y. Ueda, Synchronization of Kuramoto oscillators with time-delayed interactions and phase lag effect, J. Differ. Equ., 268 (2020), 7897–7939. https://doi.org/10.1016/j.jde.2019.11.090 doi: 10.1016/j.jde.2019.11.090
|
| [21] |
S. Kim, S. H. Park, C. S. Ryu, Multistability in coupled oscillator systems with time delay, Phys. Rev. Lett., 79 (1997), 2911. https://doi.org/10.1103/PhysRevLett.79.2911 doi: 10.1103/PhysRevLett.79.2911
|
| [22] | Y. Kuramoto, Self-entrainment of a population of coupled non-linear oscillators, in International Symposium on Mathematical Problems in Theoretical Physics, Berlin, Heidelberg: Springer, 30 (1975), 420–422. https://doi.org/10.1007/BFb0013365 |
| [23] | Y. Kuramoto, Chemical turbulence, in Chemical Oscillations, Waves and Turbulence, Berlin: Springer-Verlag, 19 1984. https://doi.org/10.1007/978-3-642-69689-3-7 |
| [24] |
V. V. Klinshov, A. A. Zlobin, Kuramoto Model with Delay: The Role of the Frequency Distribution, Mathematics, 11 (2023), 2325. https://doi.org/10.3390/math11102325 doi: 10.3390/math11102325
|
| [25] |
Z. Li, S. Y. Ha, Uniqueness and well-ordering of emergent phase-locked states for the Kuramoto model with frustration and inertia, Math. Models Methods Appl. Sci., 26 (2016), 357–382. https://doi.org/10.1142/S0218202516400054 doi: 10.1142/S0218202516400054
|
| [26] |
Z. Lin, B. Francis, M. Maggiore, State agreement for continuous-time coupled nonlinear systems, SIAM J. Control Optim., 46 (2007), 288–307. https://doi.org/10.1137/050626405 doi: 10.1137/050626405
|
| [27] |
E. Montbrió, D. Pazó, J. Schmidt, Time delay in the Kuramoto model with bimodal frequency distribution, Phys. Rev. E, 74 (2006), 056201. https://doi.org/10.1103/PhysRevE.74.056201 doi: 10.1103/PhysRevE.74.056201
|
| [28] |
E. Niebur, H. G. Schuster, D. M. Kammen, Collective frequencies and metastability in networks of limit-cycle oscillators with time delay, Phys. Rev. Lett., 67 (1991), 2753. https://doi.org/10.1103/PhysRevLett.67.2753 doi: 10.1103/PhysRevLett.67.2753
|
| [29] |
J. Parrish, L. Edelstein-Keshet, Complexity, pattern, and evolutionary trade-offs in animal aggregation, Science, 284 (1999), 99–101. https://doi.org/10.1126/science.284.5411.99 doi: 10.1126/science.284.5411.99
|
| [30] |
L. Perea, P. Elosegui, G. Gómez, Extension of the Cucker–Smale control law to space flight formations, J. Guid. Control Dyn., 32 (2009), 527–537. https://doi.org/10.2514/1.36269 doi: 10.2514/1.36269
|
| [31] |
A. Pluchino, V. Latora, A. Rapisarda, Changing opinions in a changing world: a new perspective in sociophysics, Int. J. Mod. Phys. C, 16 (2005), 515–531. https://doi.org/10.1142/S0129183105007261 doi: 10.1142/S0129183105007261
|
| [32] |
M. G. Rosenblum, A. Pikovsky, Delayed feedback control of collective synchrony: an approach to suppression of pathological brain rhythms, Phys. Rev. E, 70 (2004), 041904. https://doi.org/10.1103/PhysRevE.70.041904 doi: 10.1103/PhysRevE.70.041904
|
| [33] |
H. Sakaguchi, Y. Kuramoto, A soluble active rotator model showing phase transitions via mutual entrainment, Prog. Theor. Phys., 76 (1986), 576–581. https://doi.org/10.1143/PTP.76.576 doi: 10.1143/PTP.76.576
|
| [34] |
G. S. Schmidt, A. Papachristodoulou, U. Munz, F. Allgöwer, Frequency synchronization and phase agreement in Kuramoto oscillator networks with delays, Automatica, 48 (2012), 3008–3017. https://doi.org/10.1016/j.automatica.2012.08.013 doi: 10.1016/j.automatica.2012.08.013
|
| [35] |
H. G. Schuster, P. Wagner, Mutual entrainment of two limit cycle oscillators with time delayed coupling, Prog. Theor. Phys., 81 (1989), 939–945. https://doi.org/10.1143/PTP.81.939 doi: 10.1143/PTP.81.939
|
| [36] |
A. T. Winfree, Biological rhythms and the behavior of populations of coupled oscillators, J. Theoret. Biol., 16 (1967), 15–42. https://doi.org/10.1016/0022-5193(67)90051-3 doi: 10.1016/0022-5193(67)90051-3
|
| [37] |
M. K. S. Yeung, S. H. Strogatz, Time delay in the Kuramoto model of coupled oscillators, Phys. Rev. Lett., 82 (1999), 648–651. https://doi.org/10.1103/PhysRevLett.82.648 doi: 10.1103/PhysRevLett.82.648
|
| [38] |
X. Zhang, T. Zhu, Emergence of synchronization Kuramoto model with general digraph, Discrete Contin. Dyn. Syst. Ser. B, 28 (2023), 2335–2390. https://doi.org/10.3934/dcdsb.2022172 doi: 10.3934/dcdsb.2022172
|
| [39] |
Z. G. Zheng, Frustration effect on synchronization and chaos in coupled oscillators, Chinese Phys., 10 (2001), 703–707. http://dx.doi.org/10.1088/1009-1963/10/8/306 doi: 10.1088/1009-1963/10/8/306
|
Figures(6)
Tingting Zhu. Synchronization of the generalized Kuramoto model with time delay and frustration[J]. Networks and Heterogeneous Media, 2023, 18(4): 1772-1798. doi: 10.3934/nhm.2023077







 DownLoad:
DownLoad: