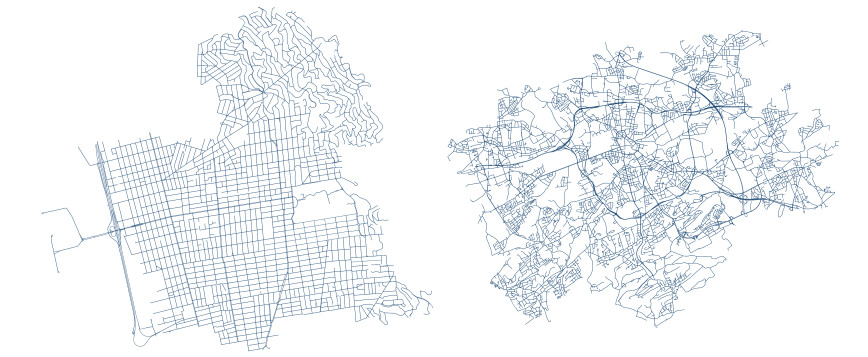
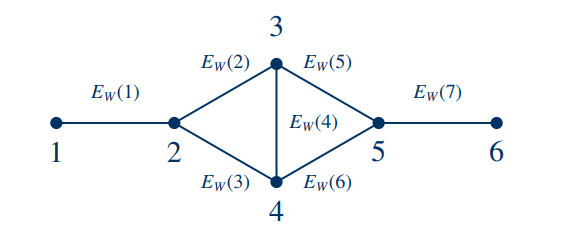
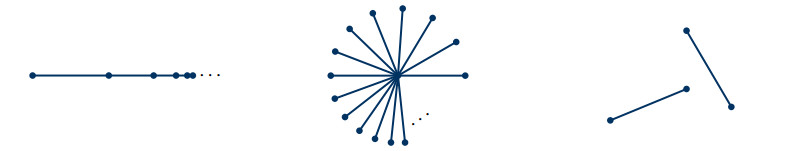
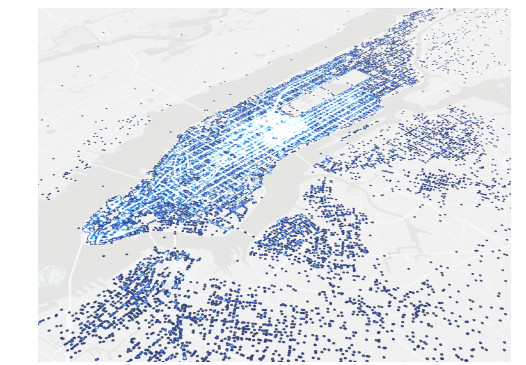
In dynamical systems on networks, Kirchhoff's first law describes the local conservation of a quantity across edges. Predominantly, Kirchhoff's first law has been conceived as a phenomenological law of continuum physics. We establish its algebraic form as a property that is inherited from fundamental axioms of a network's geometry, instead of a law observed in physical nature. To this end, we extend calculus to networks, modeled as abstract metric spaces, and derive Kirchhoff's first law for hyperbolic conservation laws. In particular, our results show that hyperbolic conservation laws on networks can be stated without explicit Kirchhoff-type boundary conditions.
Citation: Alexandre M. Bayen, Alexander Keimer, Nils Müller. A proof of Kirchhoff's first law for hyperbolic conservation laws on networks[J]. Networks and Heterogeneous Media, 2023, 18(4): 1799-1819. doi: 10.3934/nhm.2023078
In dynamical systems on networks, Kirchhoff's first law describes the local conservation of a quantity across edges. Predominantly, Kirchhoff's first law has been conceived as a phenomenological law of continuum physics. We establish its algebraic form as a property that is inherited from fundamental axioms of a network's geometry, instead of a law observed in physical nature. To this end, we extend calculus to networks, modeled as abstract metric spaces, and derive Kirchhoff's first law for hyperbolic conservation laws. In particular, our results show that hyperbolic conservation laws on networks can be stated without explicit Kirchhoff-type boundary conditions.

| [1] | L. Alon, R. Band, G. Berkolaiko, Universality of nodal count distribution in large metric graphs, Exp Math, (2022), 1–35. https://doi.org/10.1080/10586458.2022.2092565 |
| [2] | G. Bastin, J. M. Coron, Exponential stability of networks of density-flow conservation laws under PI boundary control, IFAC Proceedings Volumes, 46 (2013), 221–226. https://doi.org/10.3182/20130925-3-FR-4043.00029 |
| [3] |
G. Bastin, B. Haut, J. M. Coron, B. d'Andrea-Novel, Lyapunov stability analysis of networks of scalar conservation laws, Netw. Heterog. Media, 2 (2007), 751–759. https://doi.org/10.3934/nhm.2007.2.751 doi: 10.3934/nhm.2007.2.751

|
| [4] |
A. Bayen, J. Friedrich, A. Keimer, L. Pflug, T. Veeravalli, Modeling multilane traffic with moving obstacles by nonlocal balance laws, SIAM J. Appl. Dyn. Syst., 21 (2022), 1495–1538. https://doi.org/10.1137/20M1366654 doi: 10.1137/20M1366654
|
| [5] |
A. Bayen, A. Keimer, E. Porter, M. Spinola, Time-continuous instantaneous and past memory routing on traffic networks: A mathematical analysis on the basis of the link-delay model, SIAM J. Appl. Dyn. Syst., 18 (2019), 2143–2180. https://doi.org/10.1137/19M1258980 doi: 10.1137/19M1258980
|
| [6] |
G. Boeing, Osmnx: New methods for acquiring, constructing, analyzing, and visualizing complex street networks, Comput Environ Urban Syst, 65 (2017), 126–139. https://doi.org/10.1016/j.compenvurbsys.2017.05.004 doi: 10.1016/j.compenvurbsys.2017.05.004
|
| [7] |
D. Braess, Über ein Paradoxon aus der Verkehrsplanung, Unternehmensforschung, 12 (1968), 258–268. https://doi.org/10.1007/BF01918335 doi: 10.1007/BF01918335
|
| [8] | A. Bressan, Hyperbolic Systems of Conservation Laws: The One-dimensional Cauchy Problem, Oxford: Oxford University Press, 2000. |
| [9] |
A. Bressan, S. Čanić, M. Garavello, M. Herty, B. Piccoli, Flows on networks: recent results and perspectives, EMS Surv Math SCI, 1 (2014), 47–111. https://doi.org/10.4171/emss/2 doi: 10.4171/emss/2
|
| [10] |
G. M. Coclite, M. Garavello, Vanishing viscosity for traffic on networks, SIAM J. Math. Anal., 42 (2010), 1761–1783. https://doi.org/10.1137/090771417 doi: 10.1137/090771417
|
| [11] |
G. M. Coclite, M. Garavello, B. Piccoli, Traffic flow on a road network, SIAM J. Math. Anal., 36 (2005), 1862–1886. https://doi.org/10.1137/S0036141004402683 doi: 10.1137/S0036141004402683
|
| [12] | M. Düfel, J. B. Kennedy, D. Mugnolo, M. Plümer, M. Täufer, Boundary conditions matter: On the spectrum of infinite quantum graphs, arXiv: 2207.04024, [Preprint], (2022), [cited 2023 Nov 13]. Available from: https://doi.org/10.48550/arXiv.2207.04024 |
| [13] | M. Garavello, K. Han, B. Piccoli, Models for Vehicular Traffic on Networks, Springfield Missouri: American Institute of Mathematical Sciences, 2016. |
| [14] |
M. Garavello, B. Piccoli, Conservation laws on complex networks, Ann. Inst. Henri Poincare (C) Anal. Non Lineaire, 26 (2009), 1925–1951. https://doi.org/10.1016/j.anihpc.2009.04.001 doi: 10.1016/j.anihpc.2009.04.001
|
| [15] |
F. R. Guarguaglini, R. Natalini, Global smooth solutions for a hyperbolic chemotaxis model on a network, SIAM J. Math. Anal., 47 (2015), 4652–4671. https://doi.org/10.1137/140997099 doi: 10.1137/140997099
|
| [16] |
M. Gugat, M. Herty, A. Klar, G. Leugering, Optimal control for traffic flow networks, J Optim Theory Appl, 126 (2005), 589–616. https://doi.org/10.1007/s10957-005-5499-z doi: 10.1007/s10957-005-5499-z
|
| [17] | M. Gugat, Nodal control of conservation laws on networks, In: J. Cagnol, J. P. Zolesio (Eds.) Control and boundary analysis, Boca Raton: CRC Press, 2005, 221–236. |
| [18] |
M. Gugat, A. Keimer, G. Leugering, Z. Wang, Analysis of a system of nonlocal conservation laws for multi-commodity flow on networks, Netw. Heterog. Media, 10 (2015), 749–758. https://doi.org/10.3934/nhm.2015.10.749 doi: 10.3934/nhm.2015.10.749
|
| [19] |
H. Holden, N. H. Risebro, A mathematical model of traffic flow on a network of roads, SIAM J. Math. Anal., 26 (1995), 999–1017. https://doi.org/10.1137/S0036141093243289 doi: 10.1137/S0036141093243289
|
| [20] | J. Jost, X. Li-Jost, Calculus of Variations, Cambridge: Cambridge University Press, 1998. |
| [21] | G. Kirchhoff, Über die Auflösung der Gleichungen, auf welche man bei der Untersuchung der linearen Vertheilung galvanischer Ströme geführt wird, Annalen der Physik, 148 (1847), 497–508. |
| [22] |
T. Kottos, U. Smilansky, Periodic orbit theory and spectral statistics for quantum graphs, Ann Phys (N Y), 274 (1999), 76–124. https://doi.org/10.1006/aphy.1999.5904 doi: 10.1006/aphy.1999.5904
|
| [23] |
P. Kuchment, Quantum graphs: I. Some basic structures, Waves in Random Media, 14 (2003), S107. https://doi.org/10.1088/0959-7174/14/1/014 doi: 10.1088/0959-7174/14/1/014
|
| [24] |
N. Laurent-Brouty, A. Keimer, P. Goatin, A. M. Bayen, A macroscopic traffic flow model with finite buffers on networks: well-posedness by means of hamilton–jacobi equations, Commun Math Sci, 18 (2020), 1569–1604. https://dx.doi.org/10.4310/CMS.2020.v18.n6.a4 doi: 10.4310/CMS.2020.v18.n6.a4
|
| [25] | D. Mugnolo, Semigroup Methods for Evolution Equations on Networks, Cham: Springer, 2014. |
| [26] | D. Mugnolo, What is actually a metric graph? arXiv: 1912.07549 [Preprint], (2022), [cited 2023 Nov 13]. Available from: https://doi.org/10.48550/arXiv.1912.07549 |
| [27] |
D. Mugnolo, S. Romanelli, Dynamic and generalized wentzell node conditions for network equations, Math. Methods Appl. Sci, 30 (2007), 681–706. https://doi.org/10.1002/mma.805 doi: 10.1002/mma.805
|
| [28] |
L. O. Müller, P. J. Blanco, A high order approximation of hyperbolic conservation laws in networks: Application to one-dimensional blood flow, J. Comput. Phys., 300 (2015), 423–437. https://doi.org/10.1016/j.jcp.2015.07.056 doi: 10.1016/j.jcp.2015.07.056
|
| [29] |
M. Musch, U. S. Fjordholm, N. H. Risebro, Well-posedness theory for nonlinear scalar conservation laws on networks, Netw. Heterog. Media, 17 (2022), 101–128. https://doi.org/10.3934/nhm.2021025 doi: 10.3934/nhm.2021025
|
| [30] | OpenStreetMap Foundation board, OpenStreetMap contributors, Available from: https://planet.osm.org. |
| [31] |
L. Pauling, The diamagnetic anisotropy of aromatic molecules, J. Chem. Phys., 4 (1936), 673–677. https://doi.org/10.1063/1.1749766 doi: 10.1063/1.1749766
|
| [32] |
J. R. Platt, Classification of spectra of cata‐condensed hydrocarbons, J. Chem. Phys., 17 (1949), 484–495. https://doi.org/10.1063/1.1747293 doi: 10.1063/1.1747293
|
| [33] |
M. Richardson, N. Balazs, On the network model of molecules and solids, Ann Phys (N Y), 73 (1972), 308–325. https://doi.org/10.1016/0003-4916(72)90285-0 doi: 10.1016/0003-4916(72)90285-0
|
| [34] |
C. Wheatstone, XIII. The Bakerian lecture.–An account of several new instruments and processes for determining the constants of a voltaic circuit, Philos. Trans. R. Soc. London, 133 (1843), 303–327. https://doi.org/10.1098/rstl.1843.0014 doi: 10.1098/rstl.1843.0014
|
Figures(6)
Alexandre M. Bayen, Alexander Keimer, Nils Müller. A proof of Kirchhoff's first law for hyperbolic conservation laws on networks[J]. Networks and Heterogeneous Media, 2023, 18(4): 1799-1819. doi: 10.3934/nhm.2023078







 DownLoad:
DownLoad: