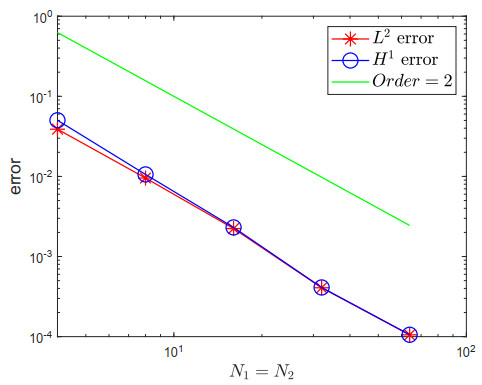
A two-dimensional multi-term time fractional diffusion equation $ {D}_{t}^{\mathit{\boldsymbol{\alpha}}} u(x, y, t)- \Delta u(x, y, t) = f(x, y, t) $ is considered in this paper, where $ {D}_{t}^{\mathit{\boldsymbol{\alpha}}} $ is the multi-term time Caputo fractional derivative. To solve the equation numerically, L1 discretisation to each fractional derivative is used on a graded temporal mesh, together with a standard finite difference method for the spatial derivatives on a uniform spatial mesh. We provide a rigorous stability and convergence analysis of a fully discrete L1-ADI scheme for solving the multi-term time fractional diffusion problem. Numerical results show that the error estimate is sharp.
Citation: Kexin Li, Hu Chen, Shusen Xie. Error estimate of L1-ADI scheme for two-dimensional multi-term time fractional diffusion equation[J]. Networks and Heterogeneous Media, 2023, 18(4): 1454-1470. doi: 10.3934/nhm.2023064
A two-dimensional multi-term time fractional diffusion equation $ {D}_{t}^{\mathit{\boldsymbol{\alpha}}} u(x, y, t)- \Delta u(x, y, t) = f(x, y, t) $ is considered in this paper, where $ {D}_{t}^{\mathit{\boldsymbol{\alpha}}} $ is the multi-term time Caputo fractional derivative. To solve the equation numerically, L1 discretisation to each fractional derivative is used on a graded temporal mesh, together with a standard finite difference method for the spatial derivatives on a uniform spatial mesh. We provide a rigorous stability and convergence analysis of a fully discrete L1-ADI scheme for solving the multi-term time fractional diffusion problem. Numerical results show that the error estimate is sharp.

| [1] |
D. Cao, H. Chen, Pointwise-in-time error estimate of an ADI scheme for two-dimensional multi-term subdiffusion equation, J. Appl. Math. Comput., 69 (2023), 707–729. https://doi.org/10.1007/s12190-022-01759-2 doi: 10.1007/s12190-022-01759-2

|
| [2] |
H. Chen, D. Xu, Y. Peng, A second order BDF alternating direction implicit difference scheme for the two-dimensional fractional evolution equation, Appl. Math. Model., 41 (2017), 54–67. https://doi.org/10.1016/j.apm.2016.05.047 doi: 10.1016/j.apm.2016.05.047
|
| [3] |
H. Chen, X. Hu, J. Ren, T. Sun, Y. Tang, L1 scheme on graded mesh for the linearized time fractional KdV equation with initial singularity, Int. J. Mod. Sim. Sci. Comp., 10 (2019), 1941006. https://doi.org/10.1142/S179396231941006X doi: 10.1142/S179396231941006X
|
| [4] | P. A. Farrell, A. F. Hegarty, J. J. H. Miller, E. O'Riordan, G. I. Shishkin, Robust computational techniques for boundary layers, Boca Raton: Chapman & Hall/CRC, 2000. |
| [5] |
X. M. Gu, H. W. Sun, Y. Zhang, Y. L. Zhao, Fast implicit difference schemes for time-space fractional diffusion equations with the integral fractional Laplacian, Math. Methods Appl. Sci., 44 (2021), 441–463. https://doi.org/10.1002/mma.6746 doi: 10.1002/mma.6746
|
| [6] |
T. Guo, O. Nikan, Z. Avazzadeh, W. Qiu, Efficient alternating direction implicit numerical approaches for multi-dimensional distributed-order fractional integro differential problems, Comput. Appl. Math., 41 (2022), 236. https://doi.org/10.1007/s40314-022-01934-y doi: 10.1007/s40314-022-01934-y
|
| [7] | R. Hilfer, Y. Luchko, Z. Tomovski, Operational method for the solution of fractional differential equations with generalized Riemann-Liouville fractional derivatives, Fract. Calc. Appl. Anal., 12 (2009), 299–318. |
| [8] |
C. Huang, H. Chen, N. An, $\beta $-robust superconvergent analysis of a finite element method for the distributed order time-fractional diffusion equation, J. Sci. Comput., 90 (2022), 44. https://doi.org/10.1007/s10915-021-01726-2 doi: 10.1007/s10915-021-01726-2
|
| [9] |
C. Huang, X. Liu, X. Meng, M. Stynes, Error analysis of a finite difference method on graded meshes for a multiterm time-fractional initial-boundary value problem, Comput. Methods Appl. Math., 20 (2020), 815–825. https://doi.org/10.1515/cmam-2019-0042 doi: 10.1515/cmam-2019-0042
|
| [10] |
C. Huang, M. Stynes, H. Chen, An $\alpha$-robust finite element method for a multi-term time-fractional diffusion problem, J. Comput. Appl. Math., 389 (2021), 113334. https://doi.org/10.1016/j.cam.2020.113334 doi: 10.1016/j.cam.2020.113334
|
| [11] |
J. Huang, J. Zhang, S. Arshad, Y. Tang, A numerical method for two-dimensional multi-term time-space fractional nonlinear diffusion-wave equations, Appl. Numer. Math., 159 (2021), 159–173. https://doi.org/10.1016/j.apnum.2020.09.003 doi: 10.1016/j.apnum.2020.09.003
|
| [12] | M. Hussain, S. Haq, Weighted meshless spectral method for the solutions of multi-term time fractional advection-diffusion problems arising in heat and mass transfer, Int J Heat Mass Tran, 129 (2019), 1305–1316. |
| [13] |
B. Li, T. Wang, X. Xie, Analysis of the L1 scheme for fractional wave equations with nonsmooth data, Comput. Math. Appl., 90 (2021), 1–12. https://doi.org/10.1016/j.camwa.2021.03.006 doi: 10.1016/j.camwa.2021.03.006
|
| [14] |
L. Li, D. Xu, M. Luo, Alternating direction implicit Galerkin finite element method for the two-dimensional fractional diffusion-wave equation, J. Comput. Phys., 255 (2013), 471–485. https://doi.org/10.1016/j.jcp.2013.08.031 doi: 10.1016/j.jcp.2013.08.031
|
| [15] |
Z. Li, Y. Liu, M. Yamamoto, Initial-boundary value problems for multi-term time-fractional diffusion equations with positive constant coefficients, Appl. Math. Comput., 257 (2015), 381–397. https://doi.org/10.1016/j.amc.2014.11.073 doi: 10.1016/j.amc.2014.11.073
|
| [16] |
Y. Lin, C. Xu, Finite difference/spectral approximations for the time-fractional diffusion equation, J. Comput. Phys., 225 (2007), 1533–1552. https://doi.org/10.1016/j.jcp.2007.02.001 doi: 10.1016/j.jcp.2007.02.001
|
| [17] |
Y. Luchko, Initial-boundary problems for the generalized multi-term time-fractional diffusion equation, J. Math. Anal. Appl., 374 (2011), 538–548. https://doi.org/10.1016/j.jmaa.2010.08.048 doi: 10.1016/j.jmaa.2010.08.048
|
| [18] | D. W. Peaceman, H. H. Rachford Jr., The numerical solution of parabolic and elliptic differential equations, J. Soc. Indust. Appl. Math., 3 (1955), 28–41. |
| [19] |
L. Qiao, J. Guo, W. Qiu, Fast BDF2 ADI methods for the multi-dimensional tempered fractional integrodifferential equation of parabolic type, Comput. Math. Appl., 123 (2022), 89–104. https://doi.org/10.1016/j.camwa.2022.08.014 doi: 10.1016/j.camwa.2022.08.014
|
| [20] |
L. Qiao, W. Qiu, D. Xu, Error analysis of fast L1 ADI finite difference/compact difference schemes for the fractional telegraph equation in three dimensions, Math. Comput. Simulation, 205 (2023), 205–231. https://doi.org/10.1016/j.matcom.2022.10.001 doi: 10.1016/j.matcom.2022.10.001
|
| [21] |
L. Qiao, D. Xu, Compact alternating direction implicit scheme for integro-differential equations of parabolic type, J. Sci. Comput., 76 (2018), 565–582. https://doi.org/10.1007/s10915-017-0630-5 doi: 10.1007/s10915-017-0630-5
|
| [22] |
S. Qin, F. Liu, I. Turner, V. Vegh, Q. Yu, Q. Yang, Multi-term time-fractional Bloch equations and application in magnetic resonance imaging, J. Comput. Appl. Math., 319 (2017), 308–319. https://doi.org/10.1016/j.cam.2017.01.018 doi: 10.1016/j.cam.2017.01.018
|
| [23] |
W. Qiu, D. Xu, H. Chen, J. Guo, An alternating direction implicit Galerkin finite element method for the distributed-order time-fractional mobile-immobile equation in two dimensions, Comput. Math. Appl., 80 (2020), 3156–3172. https://doi.org/10.1016/j.camwa.2020.11.003 doi: 10.1016/j.camwa.2020.11.003
|
| [24] |
J. Y Shen, Z. Z. Sun, R. Du, Fast finite difference schemes for time-fractional diffusion equations with a weak singularity at initial time, East Asian J. Appl. Math., 8 (2018), 834–858. https://doi.org/10.4208/eajam.010418.020718 doi: 10.4208/eajam.010418.020718
|
| [25] |
E. Sousa, C. Li, A weighted finite difference method for the fractional diffusion equation based on the Riemann-Liouville derivative, Appl. Numer. Math., 90 (2015), 22–37. https://doi.org/10.1016/j.apnum.2014.11.007 doi: 10.1016/j.apnum.2014.11.007
|
| [26] |
V. Srivastava, K. N. Rai, A multi-term fractional diffusion equation for oxygen delivery through a capillary to tissues, Math. Comput. Modelling, 51 (2010), 616–624. https://doi.org/10.1016/j.mcm.2009.11.002 doi: 10.1016/j.mcm.2009.11.002
|
| [27] | M. Stynes, A survey of the L1 scheme in the discretisation of time-fractional problems, Numer. Math. Theory Methods Appl., 15 (2022), 1173–1192. |
| [28] |
M. Stynes, E. O'Riordan, J. L. Gracia, Error analysis of a finite difference method on graded meshes for a time-fractional diffusion equation, SIAM J. Numer. Anal., 55 (2017), 1057–1079. https://doi.org/10.1137/16M1082329 doi: 10.1137/16M1082329
|
| [29] |
Y. Wang, H. Chen, T. Sun, $\alpha$-robust $H^1$-norm convergence analysis of ADI scheme for two-dimensional time-fractional diffusion equation, Appl. Numer. Math., 168 (2021), 75–83. https://doi.org/10.1016/j.apnum.2021.05.025 doi: 10.1016/j.apnum.2021.05.025
|
| [30] |
Y. Yan, M. Khan, N. J. Ford, An analysis of the modified L1 scheme for time-fractional partial differential equations with nonsmooth data, SIAM J. Numer. Anal., 56 (2018), 210–227. https://doi.org/10.1137/16M1094257 doi: 10.1137/16M1094257
|
| [31] |
Y. N. Zhang, Z. Z. Sun, Alternating direction implicit schemes for the two-dimensional fractional sub-diffusion equation, J. Comput. Phys., 230 (2011), 8713–8728. https://doi.org/10.1016/j.jcp.2011.08.020 doi: 10.1016/j.jcp.2011.08.020
|
| [32] |
Y. L. Zhao, X. M. Gu, A. Ostermann, A preconditioning technique for an all-at-once system from Volterra subdiffusion equations with graded time steps, J. Sci. Comput., 88 (2021), 11. https://doi.org/10.1007/s10915-021-01527-7 doi: 10.1007/s10915-021-01527-7
|
Figures(1) / Tables(15)
Kexin Li, Hu Chen, Shusen Xie. Error estimate of L1-ADI scheme for two-dimensional multi-term time fractional diffusion equation[J]. Networks and Heterogeneous Media, 2023, 18(4): 1454-1470. doi: 10.3934/nhm.2023064







 DownLoad:
DownLoad: