Citation: Jun Chen, Gangfeng Wang, Tao Xue, Tao Li. An improved polychromatic graphs-based BOM multi-view management and version control method for complex products[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 712-726. doi: 10.3934/mbe.2021038

| [1] | P. Burcher, Bill of materials, in Wiley Encyclopedia of Management, John Wiley & Sons, Ltd, 10 (2015), 1-1. |
| [2] | Z. Wei, H. Pan, X. Wang, Study on product BOM multi-view data definition based on object-oriented technique, Period. Ocean Univ. China, 50 (2004), 667-672. |
| [3] | R. Goncalves, D. Romero, A. Grilo, Factories of the future: challenges and leading innovations in intelligent manufacturing, Int. J. Comput. Integr. Manuf., 30 (2017), 4-13. |
| [4] | X. Liu, X. Huang, Y. Ma, Q. Meng, Y. Meng, Research on xBOM for product whole life cycle, Comput. Integr. Manuf. Syst., 8 (2002), 983-987. |
| [5] |
A. O. Aydin, A. Güngör, Effective relational database approach to represent bills-of-materials, Int. J. Prod. Res., 43 (2005), 1143-1170. doi: 10.1080/00207540512331336528

|
| [6] |
X. Huang, Y. Fan, Research on BOM views and BOM view mapping model, Chin. J. Mech. Eng., 41 (2005), 101-106. doi: 10.3901/JME.2005.12.101
|
| [7] |
F. Xiang, Y. Huang, Z. Zhang, G. Jiang, Y. Zuo, Research on ECBOM modeling and energy consumption evaluation based on BOM multi-view transformation, J. Ambient Intell. Hum. Comput., 10 (2019), 953-967. doi: 10.1007/s12652-018-1053-3
|
| [8] |
C. Zhou, X. Liu, F. Xue, H. Bo, K. Li, Research on static service BOM transformation for complex products, Adv. Eng. Inf., 36 (2018), 146-162. doi: 10.1016/j.aei.2018.02.008
|
| [9] | S. Cai, D. Hu, Y. Fei, J. Xu, Research on BOM multi-view mapping method orient to aircraft in-service data management, J. Phys.: Conf. Ser., 1550 (2020), 032124. |
| [10] | N. Xia, J. Hu, W. Wang, Managing and mapping method for BOM multiple-view based on transform-table, J. Shanghai Jiaotong Univ., 42 (2008), 584-589. |
| [11] | T. Li, Y. Li, W. Sun, G. Zhao, Research and implementation of version management method for product structure information tree, Appl. Res. Comput., 27 (2010), 3829-3833. |
| [12] | H. Zhang, S. Shi, Q. Shi, PLM-oriented BOM data management and practice, Manuf. Autom., 40 (2018), 30-34. |
| [13] | F. Liu, Y. Wang, Optimization of multi-version distribution mode for directed acyclic graph structure, Comput. Eng., 37 (2011), 74-76. |
| [14] | Y. Chen, Y. Liu, J. Liu, F. He, Design of PDM multi-version management model based on polychromatic sets theory, Advanced Materials Research, 271 (2011), 28-33. |
| [15] |
L. He, X. Ming, F. Kong, H. L. Zuo, Z. H. Yao, Research on airplane BOM management based on single source of product data, Appl. Mech. Mater., 232 (2012), 184-188. doi: 10.4028/www.scientific.net/AMM.232.184
|
| [16] |
M. Abam, S. Har-Peled, New constructions of SSPDs and their applications, Comput. Geom., 45 (2012), 200-214. doi: 10.1016/j.comgeo.2011.12.003
|
| [17] | V. Pavlov, Polychromatic Sets and Graphs for CALS, Moscow, Russia: STANKIN Press, 2002. |
| [18] |
X. Gao, L. Xu, X. Wang, Y. Li, M. Yang, Y. Liu, Workflow process modelling and resource allocation based on polychromatic sets theory, Enterp. Inf. Syst., 7 (2013), 198-226. doi: 10.1080/17517575.2012.745617
|
| [19] | S. Lin, Y. Wang, L. Jia, System reliability assessment based on failure propagation processes, Complexity, 2018 (2018), 1-19. |
| [20] | J. Leng, G. Ruan, P. Jiang, K. Xu, Q. Liu, X. Zhou, et al., Blockchain-empowered sustainable manufacturing and product lifecycle management in industry 4.0: A survey, Renewable Sustainable Energy Rev., 132 (2020), 110112. |

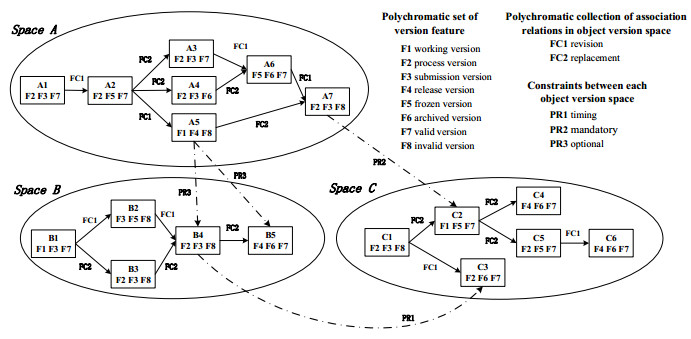
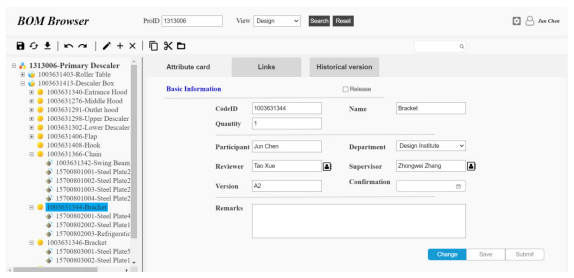
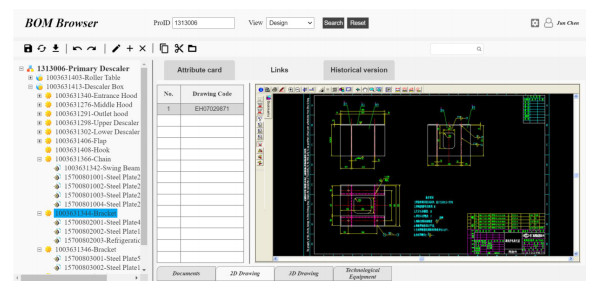
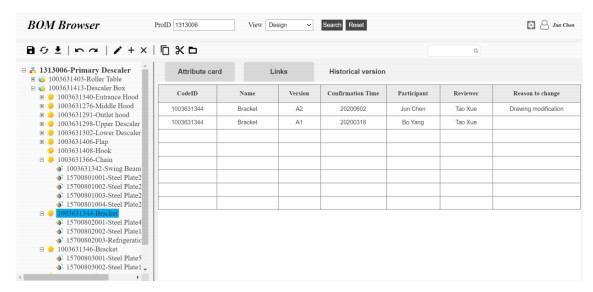
Figures(9) / Tables(4)

Jun Chen, Gangfeng Wang, Tao Xue, Tao Li. An improved polychromatic graphs-based BOM multi-view management and version control method for complex products[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 712-726. doi: 10.3934/mbe.2021038







 DownLoad:
DownLoad: