Citation: Changgui Gu, Ping Wang, Tongfeng Weng, Huijie Yang, Jos Rohling. Heterogeneity of neuronal properties determines the collective behavior of the neurons in the suprachiasmatic nucleus[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1893-1913. doi: 10.3934/mbe.2019092

| [1] | D. K. Welsh, J. S. Takahashi and S. A. Kay, Suprachiasmatic nucleus: cell autonomy and network properties, Annu. Rev. Physiol., 72 (2010), 551–577. |
| [2] | G. Zerbini, T. Kantermann and M. Merrow, Strategies to decrease social jetlag: reducing evening blue light advances sleep and melatonin, Eur. J. Neurosci., (2018). |
| [3] | A. Slomski, Melatonin Improves Sleep in Patients With Circadian Disruption, JAMA, 320 (2018), 749. |
| [4] | H. Daido, Why circadian rhythms are circadian: competitive population dynamics of biological oscillators, Phys. Rev. Lett., 87 (2001), 048101. |
| [5] | C. A. Czeisler, J. F. Duffy, T. L. Shanahan, et al., Stability, precision, and near-24-hour period of the human circadian pacemaker, Science, 284 (1999), 2177–2181. |
| [6] | J. A. Mohawk, C. B. Green and J. S. Takahashi, Central and peripheral circadian clocks in mammals, Annu. Rev. Neurosci., 35 (2012), 445–462. |
| [7] | D. K. Welsh, D. E. Logothetis, M. Meister, et al., Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms, Neuron., 14 (1995), 697–706. |
| [8] | S. Honma, W. Nakamura, T. Shirakawa, et al., Diversity in the circadian periods of single neurons of the rat suprachiasmatic nucleus depends on nuclear structure and intrinsic period, Neurosci. Lett., 358 (2004), 173–176. |
| [9] | S. M. Reppert and D. R. Weaver, Molecular analysis of mammalian circadian rhythms, Annu. Rev. Physiol., 63 (2001), 647–676. |
| [10] | E. D. Herzog, S. J. Aton, R. Numano, et al. Temporal precision in the mammalian circadian system: a reliable clock from less reliable neurons, J. Biol. Rhythms., 19 (2004), 35–46. |
| [11] | D. M. Berson, Strange vision: ganglion cells as circadian photoreceptors, Trends Neurosci., 26 (2003), 314–320. |
| [12] | S. Usui, Y. Takahashi, T. Okazaki, Range of entrainment of rat circadian rhythms to sinusoidal light-intensity cycles, Am. J. Physiol. Regul. Integr. Comp. Physiol., 278 (2000), R1148–1156. |
| [13] | U. Abraham, A. E. Granada, P. O. Westermark, et al., Coupling governs entrainment range of circadian clocks, Mol. Syst. Biol., 6 (2010): 438. |
| [14] | R. Refinetti, (2006) Circadian Physiology (2nd edn.). Boca Raton, Florida: CRC Press. |
| [15] | H. O. de la Iglesia, T. Cambras, W. J. Schwartz, et al., Forced desynchronization of dual circadian oscillators within the rat suprachiasmatic nucleus, Curr. Biol., 14 (2004), 796–800. |
| [16] | A. B. Webb, N. Angelo, J. E. Huettner, et al., Intrinsic, nondeterministic circadian rhythm generation in identified mammalian neurons, Proc. Natl. Acad. Sci. USA, 106 (2009), 16493–16498. |
| [17] | P. O. Westermark, D. K. Welsh., H. Okamura, et al., Quantification of circadian rhythms in single cells, PLoS Comput. Biol., 5 (2009), e1000580. |
| [18] | T. Hamada, J. LeSauter, J. M.Venuti, et al., Expression of Period genes: rhythmic and nonrhythmic compartments of the suprachiasmatic nucleus pacemaker, J. Neurosci., 21 (2001), 7742–7750. |
| [19] | A. B. Webb, S. R. Taylor, K. A. Thoroughman, et al., Weakly circadian cells improve resynchrony, PLoS Comput. Biol., 8 (2012), e1002787. |
| [20] | J. H. Rohling, H. T. vanderLeest, S. Michel, et al., Phase resetting of the mammalian circadian clock relies on a rapid shift of a small population of pacemaker neurons, PLoS One, 6 (2011), e25437. |
| [21] | H. Dardente, V. J. Poirel, P. Klosen, et al., Per and neuropeptide expression in the rat suprachiasmatic nuclei: compartmentalization and differential cellular induction by light, Brain Res., 958 (2002), 261–271. |
| [22] | H. T. vanderLeest, J. H. Rohling, S. Michel, et al., Phase shifting capacity of the circadian pacemaker determined by the SCN neuronal network organization, PLoS One 4 (2009), e4976. |
| [23] | M. D. Schwartz, C. Wotus, T. Liu, et al., Dissociation of circadian and light inhibition of melatonin release through forced desynchronization in the rat, Proc. Natl. Acad. Sci. U S A, 106 (2009), 17540–17545. |
| [24] | T. Noguchi, K. Watanabe, A. Ogura, et al., The clock in the dorsal suprachiasmatic nucleus runs faster than that in the ventral, Eur. J. Neurosci., 20 (2004), 3199–3202. |
| [25] | S. Yamaguchi, H. Isejima, T. Matsuo, et al., Synchronization of cellular clocks in the suprachiasmatic nucleus, Science, 302 (2003), 1408–1412. |
| [26] | H. Ohta, S. Yamazaki and D.G.McMahon, Constant light desynchronizes mammalian clock neurons, Nat. Neurosci., 8 (2005), 267–269. |
| [27] | S. J. Aton, C. S. Colwell, A. J. Harmar, et al., Vasoactive intestinal polypeptide mediates circadian rhythmicity and synchrony in mammalian clock neurons, Nat. Neurosci., 8 (2005), 476–483. |
| [28] | L. P. Morin, SCN organization reconsidered, J. Biol. Rhythms., 22 (2007), 3–13. |
| [29] | H. Albus, M. J. Vansteensel, S. Michel, et al., A GABAergic mechanism is necessary for coupling dissociable ventral and dorsal regional oscillators within the circadian clock, Curr. Biol., 15 (2005), 886–893. |
| [30] | J. A. Evans, T. L. Leise, O. Castanon-Cervantes, et al., Dynamic interactions mediated by nonredundant signaling mechanisms couple circadian clock neurons, Neuron., 80 (2013), 973–983. |
| [31] | S. R. Taylor, T. J. Wang, D. Granados-Fuentes, et al., Resynchronization Dynamics Reveal that the Ventral Entrains the Dorsal Suprachiasmatic Nucleus, J. Biol. Rhythms., 32 (2017), 35–47. |
| [32] | J. C. Leloup and A. Goldbeter, Toward a detailed computational model for the mammalian circadian clock, Proc. Natl. Acad. Sci. U S A, 100 (2003), 7051–7056. |
| [33] | H. Dardente and N. Cermakian, Molecular circadian rhythms in central and peripheral clocks in mammals, Chronobiol. Int., 24 (2007), 195–213. |
| [34] | S. Becker-Weimann, J. Wolf, H. Herzel, et al., Modeling feedback loops of the Mammalian circadian oscillator, Biophys. J., 87 (2004), 3023–3034. |
| [35] | D. Gonze, S. Bernard, C. Waltermann, et al., Spontaneous synchronization of coupled circadian oscillators, Biophys. J., 89 (2005), 120–129. |
| [36] | A. T. Winfree, (2001) The geometry of biological time. New York: Springer-Verlag. |
| [37] | S. Bernard, D. Gonze, B. Cajavec, et al., Synchronization-induced rhythmicity of circadian oscillators in the suprachiasmatic nucleus, PLoS Comput. Biol., 3 (2007), e68. |
| [38] | J. C. Locke, P. O. Westermark, A. Kramer, et al., Global parameter search reveals design principles of the mammalian circadian clock, BMC Syst. Biol., 2 (2008), 22. |
| [39] | T. Hirota and Y. Fukada, Resetting mechanism of central and peripheral circadian clocks in mammals, Zoolog. Sci., 21 (2004), 359–368. |
| [40] | C. Liu, D. R. Weaver, S. H. Strogatz, et al., Cellular construction of a circadian clock: period determination in the suprachiasmatic nuclei, Cell, 91 (1997), 855–860. |
| [41] | B. Ananthasubramaniam, E. D. Herzog and H. Herzel, Timing of neuropeptide coupling determines synchrony and entrainment in the mammalian circadian clock, PLoS Comput. Biol., 10 (2014), e1003565. |
| [42] | Z. Lu, K. Klein-Cardena, S. Lee, et al., Resynchronization of circadian oscillators and the east-west asymmetry of jet-lag,. Chaos, 26 (2016), 094811. |
| [43] | C. Schmal, J. Myung, H. Herzel, et al., A theoretical study on seasonality, Front. Neurol., 6 (2015), 94. |
| [44] | C. Vasalou, E. D. Herzog and M. A. Henson, Small-World Network Models of Intercellular Coupling Predict Enhanced Synchronization in the Suprachiasmatic Nucleus, J. Biol. Rhythms., 24 (2009), 243–254. |
| [45] | C. Vasalou and M. A. Henson, A multicellular model for differential regulation of circadian signals in the core and shell regions of the suprachiasmatic nucleus, J. Theor. Biol., 288 (2011), 44–56. |
| [46] | C. Bodenstein, M. Gosak, S. Schuster, et al., Modeling the seasonal adaptation of circadian clocks by changes in the network structure of the suprachiasmatic nucleus, PLoS Comput. Biol., 8 (2012), e1002697. |
| [47] | H. Kori and A. S. Mikhailov, Entrainment of randomly coupled oscillator networks by a pacemaker, Phys. Rev. Lett., 93 (2004), 254101. |
| [48] | H. Kori and A. S. Mikhailov, Strong effects of network architecture in the entrainment of coupled oscillator systems, Phys. Rev. E. Stat. Nonlin. Soft. Matter. Phys., 74 (2006), 066115. |
| [49] | S. Varadarajan, M. Tajiri, R. Jain, et al., Connectome of the Suprachiasmatic Nucleus: New Evidence of the Core-Shell Relationship, eNeuro., 5 (2018). |
| [50] | L. P. Morin and C. N. Allen, The circadian visual system, 2005, Brain Res. Rev., 51 (2006), 1–60. |
| [51] | J. Hannibal, M. Moller, O. P. Ottersen, et al., PACAP and glutamate are co-stored in the retinohypothalamic tract, J. Comp. Neurol., 418 (2000), 147–155. |
| [52] | D. A. Golombek, R. E. Rosenstein, Physiology of circadian entrainment, Physiol. Rev., 90 (2010), 1063–1102. |
| [53] | C. Gu, A. Ramkisoensing, Z. Liu, et al., The proportion of light-responsive neurons determines the limit cycle properties of the suprachiasmatic nucleus, J. Biol. Rhythms., 29 (2014), 16–27. |
| [54] | C. Gu, Z. Liu, W. J. Schwartz, et al, Photic desynchronization of two subgroups of circadian oscillators in a network model of the suprachiasmatic nucleus with dispersed coupling strengths, PLoS One, 7 (2012), e36900. |
| [55] | C. Gu, H. Yang, J. H. Meijer, et al., Dependence of the entrainment on the ratio of amplitudes between two subgroups in the suprachiasmatic nucleus, Phys. Rev. E., 97 (2018), 062215. |
| [56] | J. H. Meijer and W. J. Rietveld, Neurophysiology of the suprachiasmatic circadian pacemaker in rodents, Physiol. Rev., 69 (1989), 671–707. |
| [57] | C. Gu, J. Xu, Z. Liu, et al., Entrainment range of nonidentical circadian oscillators by a light-dark cycle, Phys. Rev. E. Stat. Nonlin Soft. Matter. Phys., 88 (2013), 022702. |
| [58] | C. Gu and H. Yang, The asymmetry of the entrainment range induced by the difference in intrinsic frequencies between two subgroups within the suprachiasmatic nucleus, Chaos 27 (2017), 063115. |
| [59] | C. Gu, X. Liang, H. Yang, et al., Heterogeneity induces rhythms of weakly coupled circadian neurons, Sci. Rep., 6 (2016), 21412. |
| [60] | C. Gu and H. Yang, Differences in intrinsic amplitudes of neuronal oscillators improve synchronization in the suprachiasmatic nucleus, Chaos, 27 (2017), 093108. |
| [61] | C. Gu, H. Yang and Z. Ruan, Entrainment range of the suprachiasmatic nucleus affected by the difference in the neuronal amplitudes between the light-sensitive and light-insensitive regions, Phys. Rev. E., 95 (2017), 042409. |
| [62] | C. Gu, M. Tang, J. H. Rohling, et al., The effects of non-self-sustained oscillators on the en-trainment ability of the suprachiasmatic nucleus, Sci. Rep., 6 (2016), 37661. |
| [63] | C. Gu, H. Yang and J. H. Rohling, Dissociation between two subgroups of the suprachiasmatic nucleus affected by the number of damped oscillated neurons, Phys. Rev. E., 95 (2017), 032302. |
| [64] | C. Gu, J. Wang and Z. Liu, Free-running period of neurons in the suprachiasmatic nucleus: Its dependence on the distribution of neuronal coupling strengths, Phys Rev E Stat Nonlin Soft Matter. Phys., 80 (2009), 030904. |
| [65] | C. Gu, J. H. Rohling, X. Liang, et al. Impact of dispersed coupling strength on the free running periods of circadian rhythms, Phys. Rev. E., 93 (2016), 032414. |
| [66] | C. Gu, M. Tang and H.Yang, The synchronization of neuronal oscillators determined by the directed network structure of the suprachiasmatic nucleus under different photoperiods, Sci. Rep., 6 (2016), 28878. |
| [67] | H. T. VanderLeest, T. Houben, S. Michel, et al., Seasonal encoding by the circadian pacemaker of the SCN, Curr. Biol., 17 (2007), 468–473. |
| [68] | C. Gu and H. Yang, The circadian rhythm induced by the heterogeneous network structure of the suprachiasmatic nucleus, Chaos, 26 (2016), 053112. |
| [69] | C. G. Gu, P. Wang and H. J. Yang, Entrainment range affected by the heterogeneity in the amplitude relaxation rate of suprachiasmatic nucleus neurons, Chin. Phys. B, 28 (2019), 018701. |
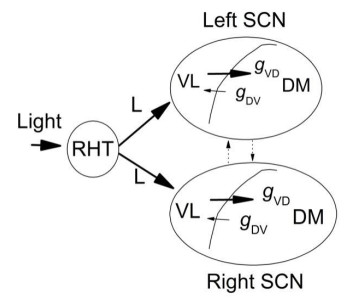
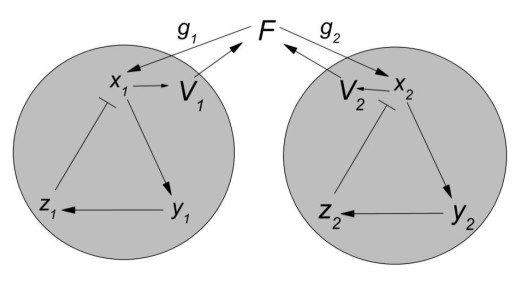
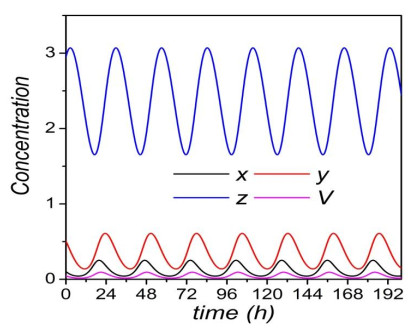
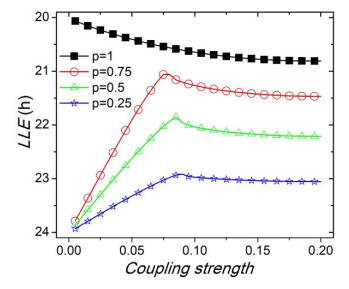
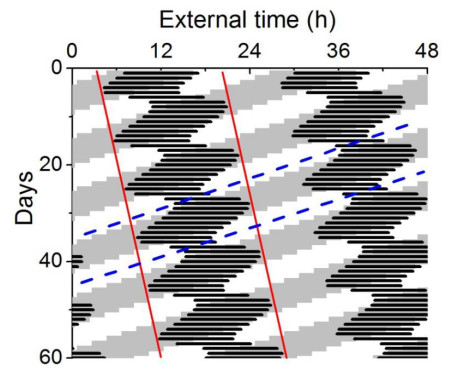
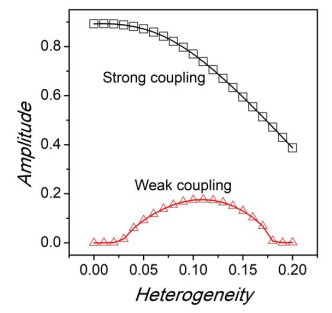
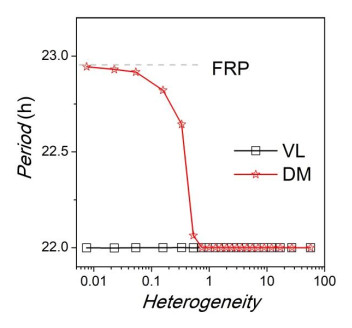
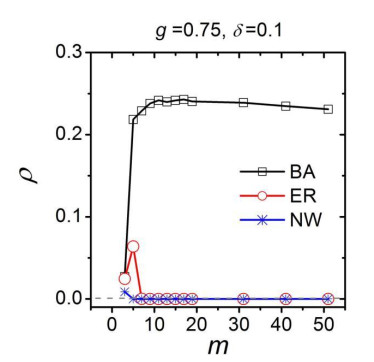
Figures(8)

Changgui Gu, Ping Wang, Tongfeng Weng, Huijie Yang, Jos Rohling. Heterogeneity of neuronal properties determines the collective behavior of the neurons in the suprachiasmatic nucleus[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1893-1913. doi: 10.3934/mbe.2019092







 DownLoad:
DownLoad: