Citation: Ana Isabel Muñoz, J. Ignacio Tello. On a mathematical model of bone marrow metastatic niche[J]. Mathematical Biosciences and Engineering, 2017, 14(1): 289-304. doi: 10.3934/mbe.2017019

| [1] | [ R. A. Adams, Sobolev Spaces Academic Press, New York-London, 1975. |
| [2] | [ A. A. Bryden,S. Islam,A. J. Freemont,J. H. Shanks,N. J. George,N. W. Clarke, Parathyroid hormone-related peptide: Expression in prostate cancer bone metastases, Prostate Cancer Prostatic Dis, 5 (2002): 59-62. |
| [3] | [ L. M. Calvi,G. B. Adams,K. W. Weibrecht,J. M. Weber,D. P. Olson,M. C. Knight,R. P. Martin,E. Schipani,P. Divieti,F. R. Bringhurst,L. A. Milner,H. M. Kronenberg,D. T. Scadden, Osteoblastic cells regulate the haematopoietic stem cell niche, Nature, 425 (2003): 841-846. |
| [4] | [ S. L. Chang, S. P. Cavnar, S. Takayama, G. D. Luker and J. J. Linderman, Cell, isoform, and environment factors shape gradients and modulate chemotaxis PLoS One 10 (2015), e0123450. |
| [5] | [ N. L. Coggins,D. Trakimas,S. L. Chang,A. Ehrlich,P. Ray,K. E. Luker,J. J. Linderman,G. D. Luker, CXCR7 controls competition for recruitment of β-arrestin 2 in cells expressing both CXCR4 and CXCR7, PLoS One, 9 (2014): 841-846. |
| [6] | [ K. Golan,O. Kollet,T. Lapidot, Dynamic cross talk between S1P and CXCL12 regulates hematopoietic stem cells migration, development and bone remodeling, Pharmaceuticals, 6 (2013): 1145-1169. |
| [7] | [ G. Innamorati,M. T. Valenti,F. Giovinazzo,L. Dalle Carbonare,M. Parenti,C. Bassi, Molecular approaches to target gpcrs in cancer therapy, Pharmaceuticals, 4 (2011): 567-589. |
| [8] | [ S. V. Komarova,R. J. Smith,S. J. Dixon,S. M. Sims,L. M. Wahlb, Mathematical model predicts a critical role for osteoclast autocrine regulation in the control of bone remodeling, Bone, 33 (2003): 206-215. |
| [9] | [ A. J. Lilly, W. E. Johnson and C. M. Bunce, The haematopoietic stem cell niche: New insights into the mechanisms regulating haematopoietic stem cell behaviour Stem Cells International, 2011 (2011), ID 274564. |
| [10] | [ A. I. Muñoz, Numerical resolution of a model of tumor growth, Mathematical Medicine and Biology, 33 (2016): 1-29. |
| [11] | [ G. O'Boyle, I. Swidenbank, H. Marshall, C. E. Barker, J. Armstrong, S. A. White, S. P. Fricker, R. Plummer, M. Wright and P. E. Lovat, Inhibition of CXCR4/CXCL12 chemotaxis in melanoma by AMD11070, Br J Cancer., 108 (2013), 1634--1640, http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3668477/ |
| [12] | [ T. Oskarsson,E. Batlle,J. Massague, Metastatic stem cells: Sources, niches, and vital pathways, Cell Stem Cell, 14 (2014): 306-321. |
| [13] | [ A. A. Rose,P. M. Siegel, Emerging therapeutic targets in breast cancer bone metastasis, Future Oncol., 6 (2010): 55-74. |
| [14] | [ M. D. Ryser,N. Nigam,S. V. Komarova, Mathematical modeling of spatio-temporal dynamics of a single bone multicellular unit, J. of Bone and Mineral Research, 24 (2009): 860-870. |
| [15] | [ J. Sceneay,M. J. Smyth,A. Möller, The pre-metastatic niche: Finding common ground, Cancer Metastasis Rev., 32 (2013): 449-464. |
| [16] | [ Y. X. Sun,J. Wang,C. E. Shelburne,D. E. Lopatin,A. M. Chinnaiyan,M. A. Rubin,K. J. Pienta,R. S. Taichman, Expression of CXCR4 and CXCL12 (SDF-1) in human prostate cancers (PCa) in vivo, Journal of Cellular Biochemistry, 89 (2003): 462-473. |
| [17] | [ R. S. Taichman,C. Cooper,E. T. Keller,K. J. Pienta,N. S. Taichman,L. K. McCauley, Use of the stromal cell-derived factor-1/CXCR4 pathway in prostate cancer metastasis to bone, Cancer Research, 62 (2002): 1832-1837. |
| [18] | [ J. I. Tello, On a mathematical model of tumor growth based on cancer stem cells, Math. Biosc. Eng., 10 (2013): 263-278. |
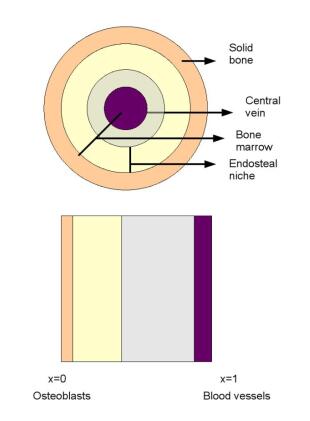
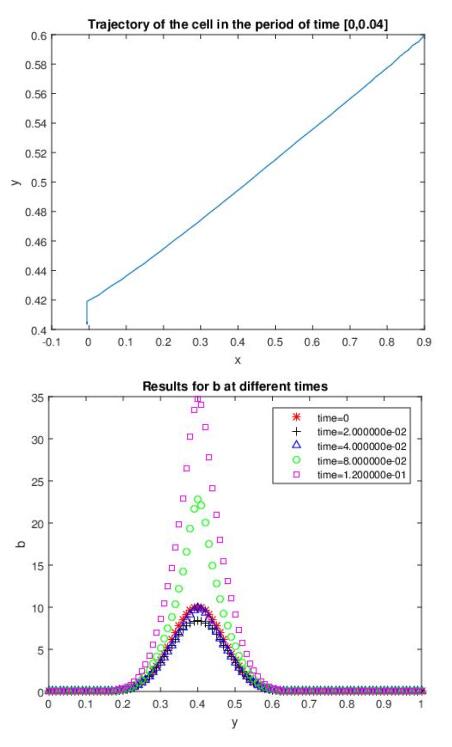
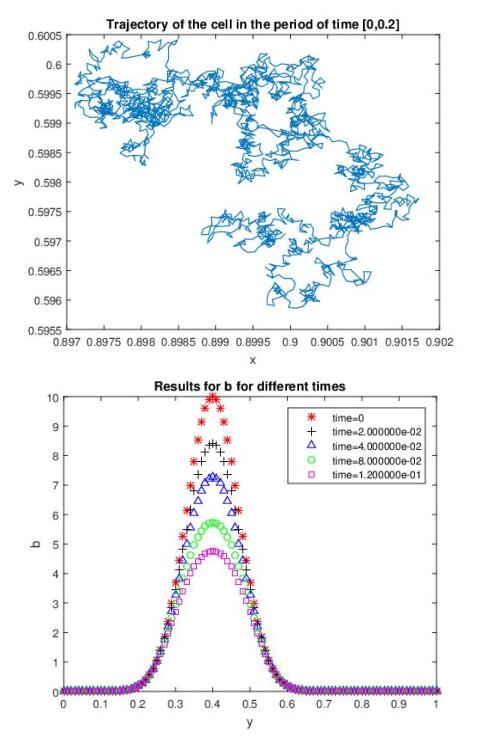
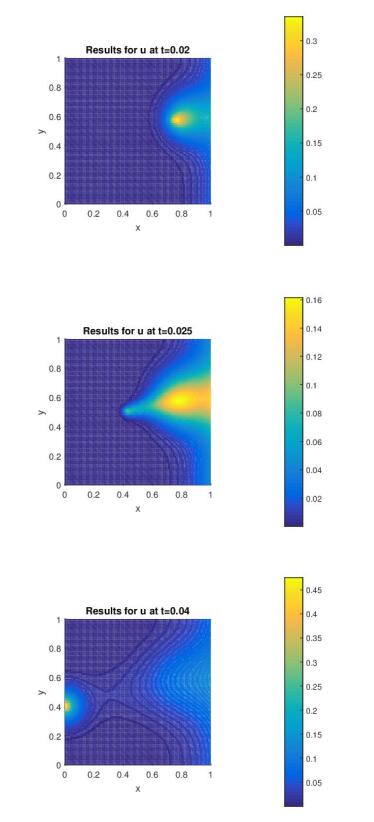
Figures(7)

Ana Isabel Muñoz, J. Ignacio Tello. On a mathematical model of bone marrow metastatic niche[J]. Mathematical Biosciences and Engineering, 2017, 14(1): 289-304. doi: 10.3934/mbe.2017019







 DownLoad:
DownLoad: