In this paper, the Holling type Ⅱ functional response extended for n different species of prey and the dynamics of interactions between one predator species and its n different classes of prey are modeled. Positivity, boundedness and permanence of all solutions of the model are proved. An ecological threshold parameter for the predator free equilibrium point of the model is established. Local stability and global stability of the predator free equilibrium point are discussed. Furthermore, we also studied that the reproduction number R0 determines whether the equilibrium points are asymptotically stable or unstable. In addition, the model was solved numerically to confirm the analytical results.
Citation: Saiwan Fatah, Arkan Mustafa, Shilan Amin. Predator and n-classes-of-prey model incorporating extended Holling type Ⅱ functional response for n different prey species[J]. AIMS Mathematics, 2023, 8(3): 5779-5788. doi: 10.3934/math.2023291
In this paper, the Holling type Ⅱ functional response extended for n different species of prey and the dynamics of interactions between one predator species and its n different classes of prey are modeled. Positivity, boundedness and permanence of all solutions of the model are proved. An ecological threshold parameter for the predator free equilibrium point of the model is established. Local stability and global stability of the predator free equilibrium point are discussed. Furthermore, we also studied that the reproduction number R0 determines whether the equilibrium points are asymptotically stable or unstable. In addition, the model was solved numerically to confirm the analytical results.

| [1] |
X. Y. Meng, N. N. Qin, H. F. Huo, Dynamics analysis of a predator-prey system with harvesting prey and disease in prey species, J. Biol. Dynam., 12 (2018), 342–374. https://doi.org/10.1080/17513758.2018.1454515 doi: 10.1080/17513758.2018.1454515

|
| [2] |
A. F. Nindjin, M. A. Aziz-Alaoui, M. Cadivel, Analysis of a predator-prey model with modified Leslie–Gower and Holling-type Ⅱ schemes with time delay, Nonlinear Anal.: Real World Appl., 7 (2006), 1104–1118. https://doi.org/10.1016/j.nonrwa.2005.10.003 doi: 10.1016/j.nonrwa.2005.10.003
|
| [3] |
K. Hassan, A. Mustafa, M. Hama, An eco-epidemiological model incorporating harvesting factors, Symmetry, 13 (2021), 2179. https://doi.org/10.3390/sym13112179 doi: 10.3390/sym13112179
|
| [4] |
I. Al-Darabsah, X. Tang, Y. Yuan, A prey-predator model with migrations and delays, Discrete Cont. Dyn. Syst.-B, 21 (2016), 737–761. https://doi.org/10.3934/dcdsb.2016.21.737 doi: 10.3934/dcdsb.2016.21.737
|
| [5] |
P. Kumar, S. Raj, Modelling and analysis of prey-predator model involving predation of mature prey using delay differential equations, Numer. Algebra, Control Optim., 12 (2022), 783–791. https://doi.org/10.3934/naco.2021035 doi: 10.3934/naco.2021035
|
| [6] |
R. K. Naji, A. N. Mustafa, The dynamics of an eco-epidemiological model with nonlinear incidence rate, J. Appl. Math., 2012 (2012), 1–24. https://doi.org/10.1155/2012/852631 doi: 10.1155/2012/852631
|
| [7] |
E, van Leeuwen, Å. Brännström, V. A. A. Jansen, U. Dieckmann, A. G. Rossberg, A generalized functional response for predators that switch between multiple prey species, J. Theor. Biol., 328 (2013), 89–98. https://doi.org/10.1016/j.jtbi.2013.02.003 doi: 10.1016/j.jtbi.2013.02.003
|
| [8] |
T. K. Kar, Stability analysis of a prey-predator model incorporating a prey refuge, Commun. Nonlinear Sci. Numer. Simul., 10 (2005), 681–691. https://doi.org/10.1016/j.cnsns.2003.08.006 doi: 10.1016/j.cnsns.2003.08.006
|
| [9] |
C. S. Holling, Some characteristics of simple types of predation and parasitism, Can. Entomol., 91 (1959), 385–398. https://doi.org/10.4039/Ent91385-7 doi: 10.4039/Ent91385-7
|
| [10] |
H. Molla, M. S. Rahman, S. Sarwardi, Dynamics of a predator-prey model with Holling type Ⅱ functional response incorporating a prey refuge depending on both the species, Int. J. Nonlinear Sci. Numer. Simul., 20 (2019), 89–104. https://doi.org/10.1515/ijnsns-2017-0224 doi: 10.1515/ijnsns-2017-0224
|
| [11] |
M. F. Elettreby, Two-prey one-predator model, Chaos, Solitons Fract., 39 (2009), 2018–2027. https://doi.org/10.1016/j.chaos.2007.06.058 doi: 10.1016/j.chaos.2007.06.058
|
| [12] |
S. Sirisubtawee, N. Khansai, A. Charoenloedmongkhon, Investigation on dynamics of an impulsive predator–prey system with generalized Holling type Ⅳ functional response and anti-predator behavior, Adv. Differ. Equ., 2021 (2021), 1–26. https://doi.org/10.1186/s13662-021-03324-w doi: 10.1186/s13662-020-03162-2
|
| [13] |
K. Das, A mathematical study of a predator-prey dynamics with disease in predator, Int. Scholarly Res. Notices, 2011 (2011), 669–684. https://doi.org/10.5402/2011/807486 doi: 10.5402/2011/807486
|
| [14] |
H. W. Hethcote, W. Wang, L. Han, Z. Ma, A predator–prey model with infected prey, Theor. Popul. Biol., 66 (2004), 259–268. https://doi.org/10.1016/j.tpb.2004.06.010 doi: 10.1016/j.tpb.2004.06.010
|
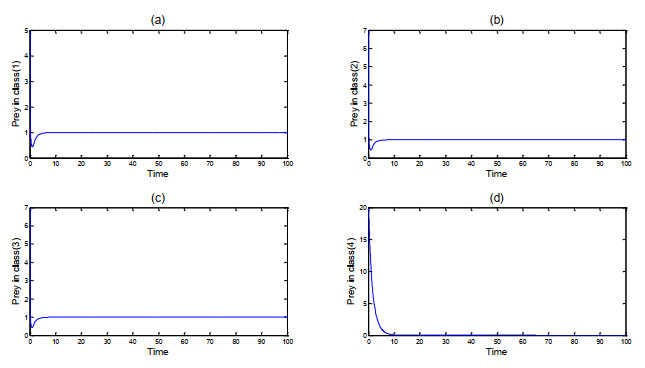

Figures(2)

Saiwan Fatah, Arkan Mustafa, Shilan Amin. Predator and n-classes-of-prey model incorporating extended Holling type Ⅱ functional response for n different prey species[J]. AIMS Mathematics, 2023, 8(3): 5779-5788. doi: 10.3934/math.2023291







 DownLoad:
DownLoad: