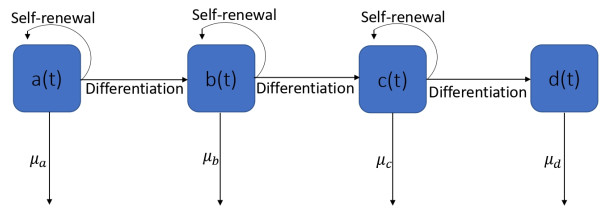
In this study, a mathematical model for the differentiation of stem cells is proposed to understand the dynamics of cell differentiation in a complex network. For this, myeloid cells, which are differentiated from stem cells, are introduced in this study. We introduce the threshold quantity $ \mathcal{R}_{0} $ to understand the population dynamics of stem cells. The local stability analysis of three equilibria, namely $ (i) $ free equilibrium points, $ (ii) $ absence of stem and progenitor cells, and $ (iii) $ endemic equilibrium points are investigated in this study. The model is first formulated in non-fractional order and after that converted into a fractional sense by utilizing the Atangana-Baleanu derivative in Caputo (ABC) sense in the form of a non-singular kernel. The model is solved by using numerical techniques. It is seen that the myeloid cell population significantly affects the stem cell population.
Citation: Ram Singh, Attiq U. Rehman, Mehedi Masud, Hesham A. Alhumyani, Shubham Mahajan, Amit K. Pandit, Praveen Agarwal. Fractional order modeling and analysis of dynamics of stem cell differentiation in complex network[J]. AIMS Mathematics, 2022, 7(4): 5175-5198. doi: 10.3934/math.2022289
In this study, a mathematical model for the differentiation of stem cells is proposed to understand the dynamics of cell differentiation in a complex network. For this, myeloid cells, which are differentiated from stem cells, are introduced in this study. We introduce the threshold quantity $ \mathcal{R}_{0} $ to understand the population dynamics of stem cells. The local stability analysis of three equilibria, namely $ (i) $ free equilibrium points, $ (ii) $ absence of stem and progenitor cells, and $ (iii) $ endemic equilibrium points are investigated in this study. The model is first formulated in non-fractional order and after that converted into a fractional sense by utilizing the Atangana-Baleanu derivative in Caputo (ABC) sense in the form of a non-singular kernel. The model is solved by using numerical techniques. It is seen that the myeloid cell population significantly affects the stem cell population.

| [1] |
A. Atangana, D. Baleanu, New fractional derivatives with the nonlocal and non-singular kernel: Theory and application to heat transfer model, J. Therm. Sci., 20 (2016), 763–769. http://dx.doi.org/10.2298/TSCI160111018A doi: 10.2298/TSCI160111018A

|
| [2] |
A. J. Wagers, J. L. Christensen, I. L. Weissman, Cell fate determination from stem cells, Gene Ther., 9 (2002), 606–612. https://doi.org/10.1038/sj.gt.3301717 doi: 10.1038/sj.gt.3301717
|
| [3] |
A. Marciniak-Czochra, T. Stiehl, W. Jaeger, A. Ho, W. Wagner, Modeling asymmetric cell division in haemtopoietic stem cells-regulation of self-renewal is essential for efficient re-population, Stem Cells Dev., 17 (2008), 1–10. https://doi.org/10.1089/scd.2008.0143 doi: 10.1089/scd.2008.0143
|
| [4] |
A. Marciniak-Czochra, T. Stiehl, W. Wagner, Modeling of replicative senescence in hematopoietic development, Aging, 1 (2009), 723–732. https://dx.doi.org/10.18632/aging.100072 doi: 10.18632/aging.100072
|
| [5] |
A. U. Rehman, R. Singh, P. Agarwal, Modeling, analysis and prediction of new variants of covid-19 and dengue co-infection on complex network, Chaos Soliton. Fract., 150 (2021), 111008. https://dx.doi.org/10.1016/j.chaos.2021.111008 doi: 10.1016/j.chaos.2021.111008
|
| [6] |
A. U. Rehman, R. Singh, T. Abdeljawad, E. Okyere, L. Guran, Modeling, analysis and numerical solution to malaria fractional model with temporary immunity and relapse, Adv. Differ. Equ., 2021 (2021), 390. https://doi.org/10.1186/s13662-021-03532-4 doi: 10.1186/s13662-021-03532-4
|
| [7] | B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walter, Molecular biology of the cell, 6 Eds., W.W. Norton & Company, Boca Raton, 2017. |
| [8] | B. E. Lawrence, P. M. Horton, Progenitor cells: Biology, characterization and potential clinical applications, Nova Science Publishers. |
| [9] |
C. Colijn, M. C. Mackey, A mathematical model of hematopoiesis–-I. Periodic chronic myelogenous leukemia, J. Theor. Biol., 237 (2005), 117–132. https://doi.org/10.1016/j.jtbi.2005.03.033 doi: 10.1016/j.jtbi.2005.03.033
|
| [10] |
C. Colijn, M. C. Mackey, A mathematical model of hematopoiesis: II. Cyclical neutropenia, J. Theor. Biol., 237 (2005), 133–146. https://doi.org/10.1016/j.jtbi.2005.03.034 doi: 10.1016/j.jtbi.2005.03.034
|
| [11] |
C. Foley, S. Bernard, M. C. Mackey, Cost-effective G-CSF therapy strategies for cyclical neutropenia: Mathematical modelling based hypotheses, J. Theor. Biol., 238 (2006), 754–763. https://doi.org/10.1016/j.jtbi.2005.06.021 doi: 10.1016/j.jtbi.2005.06.021
|
| [12] | F. R. Gantmacher, The theory of matrices, Chelsea Publishing Conmpany, New York, 1959. |
| [13] |
G. Dontu, M. Al-Hajj, W. M. Abdallah, M. F. Clarke, M. S. Wicha, Stem cells in normal breast development and breast cancer, Cell Proliferat., 36 (2003), 59–72. https://doi.org/10.1046/j.1365-2184.36.s.1.6.x doi: 10.1046/j.1365-2184.36.s.1.6.x
|
| [14] |
I. L. Weissman, Stem cells: Units of development, units of regeneration, and units of evolution, Cell, 100 (2000), 157–168. https://doi.org/10.1016/S0092-8674(00)81692-X doi: 10.1016/S0092-8674(00)81692-X
|
| [15] | I. Podlubny, Fractional differential equations: An introduction to fractional derivatives, fractional differential equations, to methods of their solution and some of their applications, Elsevier, Amsterdam, 1998. |
| [16] |
K. Dietz, J. A. P. Heesterbeek, The concept of $\mathbb{R}_{0}$ in epidemic theory, Stat. Neerl., 50 (1996), 89–110. https://doi.org/10.1111/j.1467-9574.1996.tb01482.x doi: 10.1111/j.1467-9574.1996.tb01482.x
|
| [17] |
M. Toufik, A. Atagana, New numerical approximation of fractional derivative with non-local and non-singular kernel: Application to chaotic models, Eur. Phys. J. Plus, 132 (2017), 444. https://doi.org/10.1140/epjp/i2017-11717-0 doi: 10.1140/epjp/i2017-11717-0
|
| [18] |
N. Sharma, R. Singh, J. Singh, O. Castillo, Modeling assumptions, optimal control strategies and mitigation through vaccination to zika virus, Chaos Soliton. Fract., 150 (2021), 111137. https://doi.org/10.1016/j.chaos.2021.111137 doi: 10.1016/j.chaos.2021.111137
|
| [19] |
O. Arino, M. Kimmel, Stability analysis of models of cell production system, Math. Model., 7 (1986), 1269–1300. https://doi.org/10.1016/0270-0255(86)90081-3 doi: 10.1016/0270-0255(86)90081-3
|
| [20] |
P. Agarwal, R. Singh, A. U. Rehman, Numerical solution of a hybrid mathematical model of dengue transmission with relapse and memory via Adam-Bashforth-Moulton predictor-corrector, Chaos Soliton. Fract., 143 (2021), 110564. https://doi.org/10.1016/j.chaos.2020.110564 doi: 10.1016/j.chaos.2020.110564
|
| [21] |
P. A. Beachy, S. S. Karhadkar, D. M. Berman, Tissue repair and stem cell renewal in carcinogenesis, Nature, 432 (2004), 324–331. https://doi.org/10.1038/nature03100 doi: 10.1038/nature03100
|
| [22] |
R. M. Seaberg, D. Van der Kooy, Stem and progenitor cells: The premature desertion of rigorous definitions, Trends Neurosci., 26 (2003), 125–31. https://doi.org/10.1016/S0166-2236(03)00031-6 doi: 10.1016/S0166-2236(03)00031-6
|
| [23] |
S. Bentout, Y. Chen, S. Djilali, Global dynamics of an SEIR model with two age structures and a nonlinear incidence, Acta Appl. Math., 171 (2021). https://doi.org/10.1007/s10440-020-00369-z doi: 10.1007/s10440-020-00369-z
|
| [24] |
S. Bentout, S. Djilali, B. Ghanbari, Backward, Hopf bifurcation in a heroin epidemic model with treat age, Int. J. Model. Simul. Sci. Comput., 12 (2021). https://doi.org/10.1142/S1793962321500185 doi: 10.1142/S1793962321500185
|
| [25] |
S. Djilali, L. Benahmadi, A. Tridane, K. Niri, Modeling the impact of unreported cases of the COVID-19 in the North African countries, Biology, 9 (2020), 373. https://doi.org/10.3390/biology9110373 doi: 10.3390/biology9110373
|
| [26] |
T. Sitthiwirattham, A. Zeb, S. Chasreechai, Z. Eskandari, M. Tilioua, S. Djilali, Analysis of a discrete mathematical COVID-19 model, Results Phys., 28 (2021), 104668. https://doi.org/10.1016/j.rinp.2021.104668 doi: 10.1016/j.rinp.2021.104668
|
| [27] |
T. Stiehl, A. Marciniak-Czochra, Characterization of stem cells using mathematical models of multistage cell lineages, Math. Comput. Model., 53 (2011), 1505–1517. https://doi.org/10.1016/j.mcm.2010.03.057 doi: 10.1016/j.mcm.2010.03.057
|
| [28] |
Y. Nakata, P. Getto, A. Marciniak-Czochra, T. Alarc$\acute{o}$n, Stability analysis of multi-compartment models for cell production system, J. Biol. Dynam., 6 (2012), 2–18. https://doi.org/10.1080/17513758.2011.558214 doi: 10.1080/17513758.2011.558214
|
Figures(11) / Tables(2)

Ram Singh, Attiq U. Rehman, Mehedi Masud, Hesham A. Alhumyani, Shubham Mahajan, Amit K. Pandit, Praveen Agarwal. Fractional order modeling and analysis of dynamics of stem cell differentiation in complex network[J]. AIMS Mathematics, 2022, 7(4): 5175-5198. doi: 10.3934/math.2022289







 DownLoad:
DownLoad: