In this paper, a general HIV model incorporating intracellular time delay is investigated. Taking the latent virus infection, both virus-to-cell and cell-to-cell transmissions into consideration, the model exhibits threshold dynamics with respect to the basic reproduction number $ \mathfrak{R}_0 $. If $ \mathfrak{R}_0 < 1 $, then there exists a unique infection-free equilibrium $ E_0 $, which is globally asymptotically stable. If $ \mathfrak{R}_0 > 1 $, then there exists $ E_0 $ and a globally asymptotically stable infected equilibrium $ E^* $. When $ \mathfrak{R}_0 = 1 $, $ E_0 $ is linearly neutrally stable and a forward bifurcation takes place without time delay around $ \mathfrak{R}_0 = 1 $. The theoretical results and corresponding numerical simulations show that the existence of latently infected cells and the intracellular time delay have vital effect on the global dynamics of the general virus model.
Citation: Xin Jiang. Threshold dynamics of a general delayed HIV model with double transmission modes and latent viral infection[J]. AIMS Mathematics, 2022, 7(2): 2456-2478. doi: 10.3934/math.2022138
In this paper, a general HIV model incorporating intracellular time delay is investigated. Taking the latent virus infection, both virus-to-cell and cell-to-cell transmissions into consideration, the model exhibits threshold dynamics with respect to the basic reproduction number $ \mathfrak{R}_0 $. If $ \mathfrak{R}_0 < 1 $, then there exists a unique infection-free equilibrium $ E_0 $, which is globally asymptotically stable. If $ \mathfrak{R}_0 > 1 $, then there exists $ E_0 $ and a globally asymptotically stable infected equilibrium $ E^* $. When $ \mathfrak{R}_0 = 1 $, $ E_0 $ is linearly neutrally stable and a forward bifurcation takes place without time delay around $ \mathfrak{R}_0 = 1 $. The theoretical results and corresponding numerical simulations show that the existence of latently infected cells and the intracellular time delay have vital effect on the global dynamics of the general virus model.

| [1] |
A. Alshorman, X. Wang, M. J. Meyer, L. Rong, Analysis of HIV models with two time delays, J. Biol. Dyn., 11 (2017), 40-64. doi: 10.1080/17513758.2016.1148202. doi: 10.1080/17513758.2016.1148202

|
| [2] |
S. Bonhoeffer, R. M. May, G. M. Shaw, M. A. Nowak, Virus dynamics and drug therapy, Proc. Natl. Acad. Sci. USA, 94 (1997), 6971-6976. doi: 10.1073/pnas.94.13.6971. doi: 10.1073/pnas.94.13.6971
|
| [3] |
C. Castillo-Chavez, B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361-404. doi: 10.3934/mbe.2004.1.361. doi: 10.3934/mbe.2004.1.361
|
| [4] |
P. Chen, W. Hübner, M. A. Spinelli, B. K. Chen, Predominant mode of human immunodeficiency virus transfer between T cells is mediated by sustained Env-dependent neutralization-resistant virological synapses, J. Virol., 81 (2007), 12582-12595. doi: 10.1128/JVI.00381-07. doi: 10.1128/JVI.00381-07
|
| [5] |
T. W. Chun, L. Stuyver, S. B. Mizell, L. A. Ehler, J. A. M. Mican, M. Baseler, et al., Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy, Proc. Natl. Acad. Sci., 94 (1997), 13193-13197. doi: 10.1073/pnas.94.24.13193. doi: 10.1073/pnas.94.24.13193
|
| [6] |
S. M. Ciupe, Modeling the dynamics of hepatitis B infection, immunity, and drug therapy, Immunol. Rev., 285 (2018), 38-54. doi: 10.1111/imr.12686. doi: 10.1111/imr.12686
|
| [7] |
D. S. Dimitrov, R. L. Willey, H. Sato, L. Chang, R. Blumenthal, M. A. Martin, Quantitation of human immunodeficiency virus type 1 infection kinetics, J. Virol., 67 (1993), 2182-2190. doi: 10.1128/JVI.67.4.2182-2190.1993. doi: 10.1128/JVI.67.4.2182-2190.1993
|
| [8] |
S. Gummuluru, C. M. Kinsey, M. Emerman, An in vitro rapid-turnover assay for human immunodeficiency virus type 1 replication selects for cell-to-cell spread of virua, J. Virol., 74 (2000), 10882-10891. doi: 10.1128/jvi.74.23.10882-10891.2000. doi: 10.1128/jvi.74.23.10882-10891.2000
|
| [9] | J. K. Hale, S. M. V. Lunel, Introduction to functional differential equations, New York: Springer-Verlag, 1993. doi: 10.1007/978-1-4612-4342-7. |
| [10] |
A. V. Herz, S. Bonhoeffer, R. M. Anderson, R. M. May, M. A. Nowak, Viral dynamics in vivo: Limitations on estimations on intracellular delay and virus delay, Proc. Natl. Acad. Sci. USA, 93 (1996), 7247-7251. doi: 10.1073/pnas.93.14.7247. doi: 10.1073/pnas.93.14.7247
|
| [11] |
W. Hübner, G. P. McNerney, P. Chen, B. M. Dale, R. E. Gordon, F. Y. S. Chuang, et al., Quantitative 3D video microscopy of HIV transfer across T cell virological synapses, Science, 323 (2009), 1743-1747. doi: 10.1126/science.1167525. doi: 10.1126/science.1167525
|
| [12] |
D. E. Kirschner, G. F. Webb, A mathematical model of combined drug therapy of HIV infection, Comput. Math. Methods Med., 1 (1997), 293715. doi: 10.1080/10273669708833004. doi: 10.1080/10273669708833004
|
| [13] |
A. Korobeinikov, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 876-883. doi: 10.1016/j.bulm.2004.02.001. doi: 10.1016/j.bulm.2004.02.001
|
| [14] |
T. Kuniya, Hopf bifurcation in an age-structured SIR epidemic model, Appl. Math. Lett., 92 (2019), 22-28. doi: 10.1016/j.aml.2018.12.010. doi: 10.1016/j.aml.2018.12.010
|
| [15] | Y. Kuang, Delay differential equations with applications in population dynamics, New York: Academic Press, 1993. |
| [16] |
X. Lai, X. Zou, Modelling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898-917. doi: 10.1137/130930145. doi: 10.1137/130930145
|
| [17] |
K. Lassen, Y. Han, Y. Zhou, J. Siliciano, R. F. Siliciano, The multifactorial nature of HIV-1 latency, Trends Mol. Med., 11 (2004), 525-531. doi: 10.1016/j.molmed.2004.09.006. doi: 10.1016/j.molmed.2004.09.006
|
| [18] |
J. Lin, R. Xu, X. Tian, Threshold dynamics of an HIV-1 virus model with both virus-to-cell and cell-to-cell transmissions, intracellular delay, and humoral immunity, Appl. Math. Comput., 315 (2017), 516-530. doi: 10.1016/j.amc.2017.08.004. doi: 10.1016/j.amc.2017.08.004
|
| [19] |
C. C. McCluskey, Using Lyapunov functions to construct Lyapunov functionals for delay differential equations, SIAM J. Appl. Dyn. Syst., 14 (2015), 1-24. doi: 10.1137/140971683. doi: 10.1137/140971683
|
| [20] |
C. C. McCluskey, Y. Yang, Global stability of a diffusive virus dynamics model with general incidence function and time delay, Nonlinear Anal.: Real World Appl., 25 (2015), 64-78. doi: 10.1016/j.nonrwa.2015.03.002. doi: 10.1016/j.nonrwa.2015.03.002
|
| [21] |
F. E. McKenzie, W. H. Bossert, A target for intervention in plasmodium falciparum infections, Am. J. Trop. Med. Hyg., 58 (1998), 763-767. doi: 10.4269/ajtmh.1998.58.763. doi: 10.4269/ajtmh.1998.58.763
|
| [22] |
A. S. Perelson, D. E. Kirschner, R. J. De Boer, Dynamics of HIV infection of CD4+ T cells, Math. Biosci., 114 (1993), 81-125. doi: 10.1016/0025-5564(93)90043-A. doi: 10.1016/0025-5564(93)90043-A
|
| [23] |
A. S. Perelson, A. U. Neumann, M. Markowitz, J. M. Leonard, D. D. Ho, HIV-1 dynamics in vivo: Virion clearance rate, infected cell life-span, and viral generation time, Science, 271 (1996), 1582-1586. doi: 10.1126/science.271.5255.1582. doi: 10.1126/science.271.5255.1582
|
| [24] |
D. M. Philips, The role of cell-to-cell transmission in HIV infection, AIDS, 8 (1994), 719-731. doi: 10.1097/00002030-199406000-00001. doi: 10.1097/00002030-199406000-00001
|
| [25] |
H. Pourbashash, S. S. Pilyugin, P. D. Leenheer, C. McCluskey, Global analysis of within host virus models with cell-to-cell viral transmission, Discrete Contin. Dyn. Syst. Ser. B, 19 (2014), 3341-3357. doi: 10.3934/dcdsb.2014.19.3341. doi: 10.3934/dcdsb.2014.19.3341
|
| [26] |
B. Ramratnam, S. Bonhoeffer, J. Binley, A. Hurley, L. Zhang, J. E. Mittler, et al., Rapid production and clearance of HIV-1 and hepatitis C virus assessed by large volume plasma apheresis, Lancet, 354 (1999), 1782-1785. doi: 10.1016/S0140-6736(99)02035-8. doi: 10.1016/S0140-6736(99)02035-8
|
| [27] |
L. Rong, Z. Feng, A. S. Perelson, Emergence of HIV-1 drug resistance during antiretroviral treatment, Bull. Math. Biol., 69 (2007), 2027-2060. doi: 10.1007/s11538-007-9203-3. doi: 10.1007/s11538-007-9203-3
|
| [28] |
L. Rong, A. S. Perelson, Modeling HIV persistence, the latent reservoir, and viral blips, J. Theoret. Biol., 260 (2009), 308-331. doi: 10.1016/j.jtbi.2009.06.011. doi: 10.1016/j.jtbi.2009.06.011
|
| [29] |
L. Rong, A. S. Perelson, Modeling Latently infected cell activation: Viral and latent reservoir persistence, and viral blips in HIV-infected patients on potent therapy, PLoS Comput. Biol., 5 (2009), e1000533. doi: 10.1371/journal.pcbi.1000533. doi: 10.1371/journal.pcbi.1000533
|
| [30] |
H. Sato, J. Orenstein, D. S. Dimitrov, M. A. Martin, Cell-to-cell spread of HIV-1 occurs with minutes and may not involve the participation of virus particles, Virology, 186 (1992), 712-724. doi: 10.1016/0042-6822(92)90038-q. doi: 10.1016/0042-6822(92)90038-q
|
| [31] |
Q. Sattentau, The direct passage of animal viruses between cells, Curr. Opin. Virol., 1 (2011), 396-402. doi: 10.1016/j.coviro.2011.09.004. doi: 10.1016/j.coviro.2011.09.004
|
| [32] |
H. Shu, Y. Chen, L. Wang, Impacts of the cell-free and cell-to-cell infection modes on viral dynamics, J. Dyn. Diff. Equat., 30 (2018), 1817-1836. doi: 10.1007/s10884-017-9622-2. doi: 10.1007/s10884-017-9622-2
|
| [33] |
A. Sigal, J. T. Kim, A. B. Balazs, E. Dekel, A. Mayo, R. Milo, et al., Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95-98. doi: 10.1038/nature10347. doi: 10.1038/nature10347
|
| [34] |
H. Sun, J. Wang, Dynamics of a diffusive virus model with general incidence function, cell-to-cell transmission and time delay, Comput. Math. Appl., 77 (2019), 284-301. doi: 10.1016/j.camwa.2018.09.032. doi: 10.1016/j.camwa.2018.09.032
|
| [35] |
S. Tang, Z. Teng, H. Miao, Global dynamics of a reaction-diffusion virus infection model with humoral immunity and nonlinear incidence, Comput. Math. Appl., 78 (2019), 786-806. doi: 10.1016/j.camwa.2019.03.004. doi: 10.1016/j.camwa.2019.03.004
|
| [36] |
K. Wang, W. Wang, Propagation of HBV with spatial dependence, Math. Biosci., 210 (2007), 78-95. doi: 10.1016/j.mbs.2007.05.004. doi: 10.1016/j.mbs.2007.05.004
|
| [37] |
W. Wang, X. Wang, K. Guo, W. Ma, Global analysis of a diffusive viral model with cell-to-cell infection and incubation period, Math. Meth. Appl. Sci., 43 (2020), 5963-5978. doi: 10.1002/mma.6339. doi: 10.1002/mma.6339
|
| [38] |
X. Wang, S. Tang, X. Song, L. Rong, Mathematical analysisi of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dyn., 11 (2017), 455-483. doi: 10.1080/17513758.2016.1242784. doi: 10.1080/17513758.2016.1242784
|
| [39] |
J. K. Wong, M. Hezareh, H. F. Günthard, D. V. Havlir, C. C. Ignacio, C. A. Spina, et al., Recovery of replication-competent HIV despite prolonged suppression of plasma viremia, Science, 278 (1997), 1291-1295. doi: 10.1126/science.278.5341.1291. doi: 10.1126/science.278.5341.1291
|
| [40] |
Y. Yang, L. Zou, S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183-191. doi: 10.1016/j.mbs.2015.05.001. doi: 10.1016/j.mbs.2015.05.001
|
| [41] |
X. Zhang, Z. Liu, Bifurcation analysis of an age structured HIV infection model with both virus-to-cell and cell-to-cell transmissions, Int. J. Bifurcation and Chaos, 28 (2018), 1850109. doi: 10.1142/S0218127418501092. doi: 10.1142/S0218127418501092
|
| [42] | Fact sheet-Latest statistics on the status of the AIDS epidemic, UNAIDS, 2021. Available from: https://www.unaids.org/sites/default/files/media_asset/UNAIDS_FactSheet_en.pdf. |
| [43] | FDA approval of HIV medicines, AIDSinfo, 2021. Available from: https://aidsinfo.nih.gov/understanding-hiv-aids/infographics/25/fda-approval-of-hiv-medicines. |
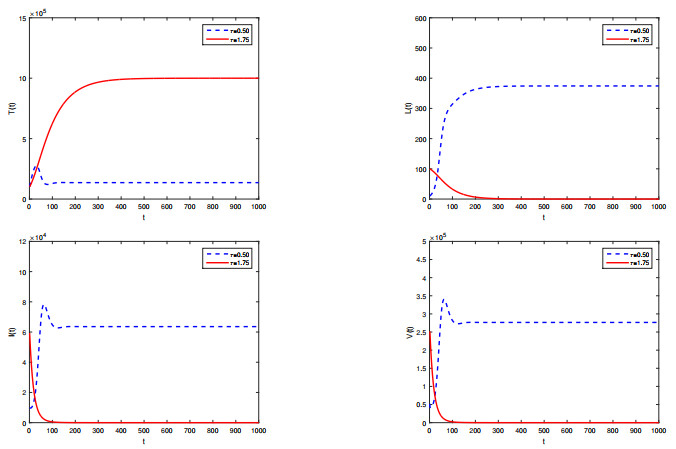
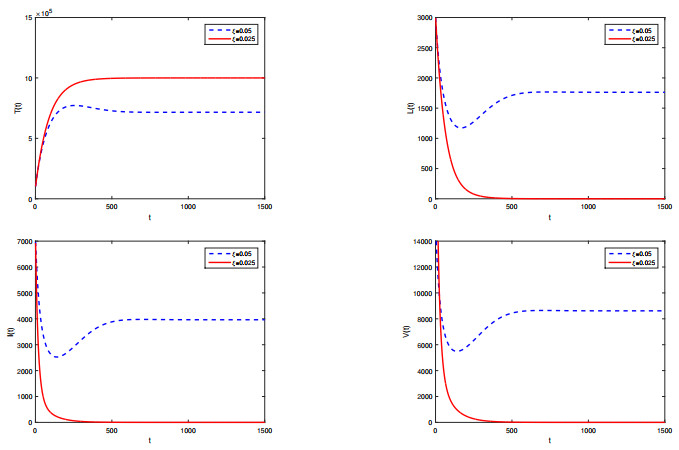
Figures(2) / Tables(1)

Xin Jiang. Threshold dynamics of a general delayed HIV model with double transmission modes and latent viral infection[J]. AIMS Mathematics, 2022, 7(2): 2456-2478. doi: 10.3934/math.2022138







 DownLoad:
DownLoad: