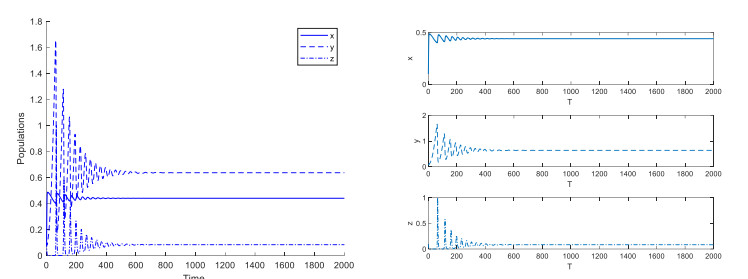
In this paper, we investigate an aquatic ecological model of microcystis aeruginosa-filter feeding fish and predatory fish model with aggregation effect of microcystis aeruginosa. Fear effect of predatory fish on filter feeding fish and harvesting effect of big fish is considered. Mathematical analysis includes two parts. The first is theoretical part, which includes proving the positive and constraining solutions of the model. Also finding equilibrium points and studying their local stability is included in this part. In addition, analyzing the local bifurcation of equilibrium points and indicating the type of bifurcation is discussed here. On the other hand, the second part contains the numerical simulation of all the theoretical results, where we compare the numerical values of the conditions obtained in the theoretical part.
Citation: Ashraf Adnan Thirthar, Salam J. Majeed, Kamal Shah, Thabet Abdeljawad. The dynamics of an aquatic ecological model with aggregation, Fear and Harvesting Effects[J]. AIMS Mathematics, 2022, 7(10): 18532-18552. doi: 10.3934/math.20221018
In this paper, we investigate an aquatic ecological model of microcystis aeruginosa-filter feeding fish and predatory fish model with aggregation effect of microcystis aeruginosa. Fear effect of predatory fish on filter feeding fish and harvesting effect of big fish is considered. Mathematical analysis includes two parts. The first is theoretical part, which includes proving the positive and constraining solutions of the model. Also finding equilibrium points and studying their local stability is included in this part. In addition, analyzing the local bifurcation of equilibrium points and indicating the type of bifurcation is discussed here. On the other hand, the second part contains the numerical simulation of all the theoretical results, where we compare the numerical values of the conditions obtained in the theoretical part.

| [1] | M. Begon, J. Harper, Townsend CR-1990: Ecology: Individuals, Populations and Communities, Oxford: Blackwell 1990. https://www.amazon.com/Ecology-Individuals-Populations-Communities-Begon/dp/0632038012 |
| [2] |
L. M. Campbell, R. E. Hecky, J. Nyaundi, R. Muggide, D. G. Dixon, Distribution and food-web transfer of mercury in Napoleon and Winam Gulfs, Lake Victoria, East Africa, J. Great Lakes Res., 29 (2003), 267–282. https://doi.org/10.1016/S0380-1330(03)70554-1 doi: 10.1016/S0380-1330(03)70554-1

|
| [3] |
J. M. Neff, S. A. Stout, D. G. Gunster, Ecological risk assessment of polycyclic aromatic hydrocarbons in sediments: Identifying sources and ecological hazard, Integr. Environ. Assess., 1 (2005), 22–33. https://doi.org/10.1897/IEAM_2004a-016.1 doi: 10.1897/IEAM_2004a-016.1
|
| [4] | E. Kerkhoven, T. Y. Gan, Development of a hydrologic scheme for use in land surface models and its application to climate change in the Athabasca River Basin. In: Cold Region Atmospheric and Hydrologic Studies, The Mackenzie GEWEX Experience, 411–433, Springer, Berlin, Heidelberg, 2008. https://doi.org/10.1007/978-3-540-75136-6_22 |
| [5] |
X. Liu, X. Lu, Y. Chen, The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: An 11-year investigation, Harmful Algae, 10 (2011), 337–343. https://doi.org/10.1016/j.hal.2010.12.002 doi: 10.1016/j.hal.2010.12.002
|
| [6] | Y. S. Zhang, H. Y. Li, F. X. Kong, Y. Yu, M. Zhang, Role of conony intercellular space in the cyanobacteria bloom-forming, Environ. Sci., 32 (2011), 1602–1607. |
| [7] |
S. Pereira, A. Zille, E. Micheletti, P. Moradas-Ferreira, R. De Philippis, P. Tamagnini, Complexity of cyanobacterial exopolysaccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly, FEMS Microbiol. Rev., 33 (2009), 917–941. https://doi.org/10.1111/j.1574-6976.2009.00183.x doi: 10.1111/j.1574-6976.2009.00183.x
|
| [8] |
Z. Yang, F. Kong, X. Shi, H. Cao, Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton, Hydrobiology, 563 (2006), 225–230. https://doi.org/10.1007/s10750-005-0008-9 doi: 10.1007/s10750-005-0008-9
|
| [9] |
H. Shen, L. Song, Comparative studies on physiological responses to phosphorus in two phenotypes of bloom-forming Microcystis, Hydrobiology, 592 (2007), 475–486. https://doi.org/10.1007/s10750-007-0794-3 doi: 10.1007/s10750-007-0794-3
|
| [10] |
H. Liu, Z. Li, M. Gao, H. Dai, Z. Liu, Dynamics of a host–parasitoid model with Allee effect for the host and parasitoid aggregation, Ecol. Complex., 6 (2009), 337–345. https://doi.org/10.1016/j.ecocom.2009.01.003. doi: 10.1016/j.ecocom.2009.01.003
|
| [11] |
M. Detto, H. C. Muller-Landau, Stabilization of species coexistence in spatial models through the aggregation-segregation effect generated by local dispersal and nonspecific local interactions, Theor. Popul. Biol., 112 (2016), 97–108. https://doi.org/10.1016/j.tpb.2016.08.008 doi: 10.1016/j.tpb.2016.08.008
|
| [12] |
X. Li, H. Yu, C. Dai, Z. Ma, Q. Wang, M. Zhao, Bifurcation analysis of a new aquatic ecological model with aggregation effect, Math. Comput. Simulat., 190 (2021), 75–96. https://doi.org/10.1016/j.matcom.2021.05.015 doi: 10.1016/j.matcom.2021.05.015
|
| [13] |
P. Xie, J. Liu, Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms: A synthesis of decades of research and application in a subtropical hypereutrophic lake, The Scientific World J., 1 (2001), 337–356. https://doi.org/10.1100/tsw.2001.67 doi: 10.1100/tsw.2001.67
|
| [14] |
U. Sommer, E. Charalampous, M. Scotti, M. Moustaka-Gouni, Big fish eat small fish: Implications for food chain length? Community Ecol., 19 (2018), 107–115. https://doi.org/10.1556/168.2018.19.2.2. doi: 10.1556/168.2018.19.2.2
|
| [15] | R. Ehrenberg, Theoretische Biologie: Vom Standpunkt der Irreversibilität des elementaren Lebensvorganges, Springer-Verlag, New York, 2013. https://link.springer.com/article/10.1007/BF00444193 |
| [16] |
H. Yu, M. Zhao, R. P. Agarwal, Stability and dynamics analysis of time delayed eutrophication ecological model based upon the Zeya reservoir, Math. Comput. Simul., 97 (2014), 53–67. https://doi.org/10.1016/j.matcom.2013.06.008 doi: 10.1016/j.matcom.2013.06.008
|
| [17] |
H. Malchow, S. Petrovskii, A. Medvinsky, Pattern formation in models of plankton dynamics, A synthesis, Oceanol. Acta, 24 (2011), 479–487. https://doi.org/10.1016/S0399-1784(01)01161-6 doi: 10.1016/S0399-1784(01)01161-6
|
| [18] |
H. Yu, M. Zhao, Q. Wang, R. P. Agarwal, A focus on long-run sustainability of an impulsive switched eutrophication controlling system based upon the Zeya reservoir, J. Franklin I., 351 (2014), 487–499. https://doi.org/10.1016/j.jfranklin.2013.08.025. doi: 10.1016/j.jfranklin.2013.08.025
|
| [19] | S. J. Majeed, R. K. Naji, A. A. Thirthar, The dynamics of an Omnivore-predator-prey model with harvesting and two different nonlinear functional responses. In: AIP Conference Proceedings (Vol. 2096, No. 1, p. 020008), April, 2019, AIP Publishing LLC, New York. |
| [20] |
X. Liu, Y. Lou, Global dynamics of a predator-prey model, J. Math. Anal. Appl., 371 (2010), 323–340. https://doi.org/10.1016/j.jmaa.2010.05.037. doi: 10.1016/j.jmaa.2010.05.037
|
| [21] |
Y. Lv, R. Yuan, Y. Pei, A prey-predator model with harvesting for fishery resource with reserve area, Appl. Math. Model., 37 (2013), 3048–3062. https://doi.org/10.1016/j.apm.2012.07.030 doi: 10.1016/j.apm.2012.07.030
|
| [22] |
F. B. Yousef, A. Yousef, C. Maji, Effects of fear in a fractional-order predator-prey system with predator density-dependent prey mortality, Chaos, Soliton. Fract., 145 (2021), 110711. https://doi.org/10.1016/j.chaos.2021.110711 doi: 10.1016/j.chaos.2021.110711
|
| [23] |
D. Mukherjee, Study of fear mechanism in predator-prey system in the presence of competitor for the prey, Ecol. Genet. Genomics, 15 (2020), 100052. https://doi.org/10.1016/j.egg.2020.100052 doi: 10.1016/j.egg.2020.100052
|
| [24] |
M. F. Danca, M. Fečkan, N. Kuznetsov, G. Chen, Rich dynamics and anticontrol of extinction in a prey–predator system, Nonlinear Dynam., 98 (2019), 1421–1445. https://doi.org/10.1007/s11071-019-05272-3 doi: 10.1007/s11071-019-05272-3
|
| [25] |
L. Y. Zanette, A. F. White, M. C. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [26] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Biol., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [27] | S. Creel, D. Christianson, Relationships between direct predation and risk effects, Trends Ecol. Evol., 23 (2008), 194–201. https://10.1016/j.tree.2007.12.004. |
| [28] |
S. L. Lima, Nonlethal effects in the ecology of predator-prey Interactions, Bioscience, 48 (1998), 25–34. https://doi.org/10.2307/1313225. doi: 10.2307/1313225
|
| [29] |
P. Panday, N. Pal, S. Samanta, J. Chattopadhyay, Stability and bifurcation analysis of a three-species food chain model with fear, Int. J. Bifurcat. Chaos, 28 (2018), 1850009. https://doi.org/10.1142/S0218127418500098 doi: 10.1142/S0218127418500098
|

















Figures(18)

Ashraf Adnan Thirthar, Salam J. Majeed, Kamal Shah, Thabet Abdeljawad. The dynamics of an aquatic ecological model with aggregation, Fear and Harvesting Effects[J]. AIMS Mathematics, 2022, 7(10): 18532-18552. doi: 10.3934/math.20221018







 DownLoad:
DownLoad: