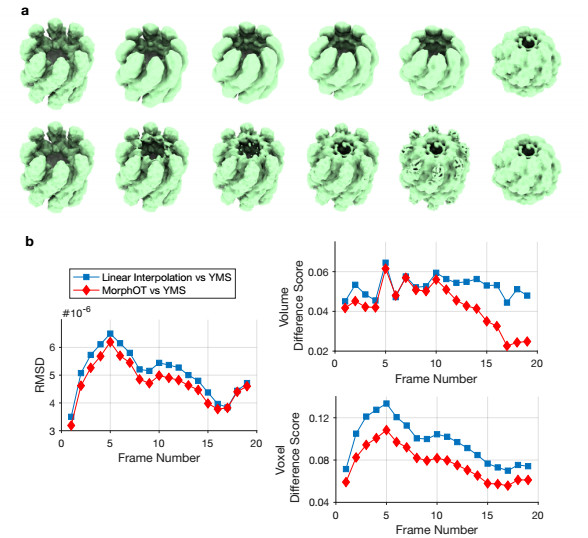
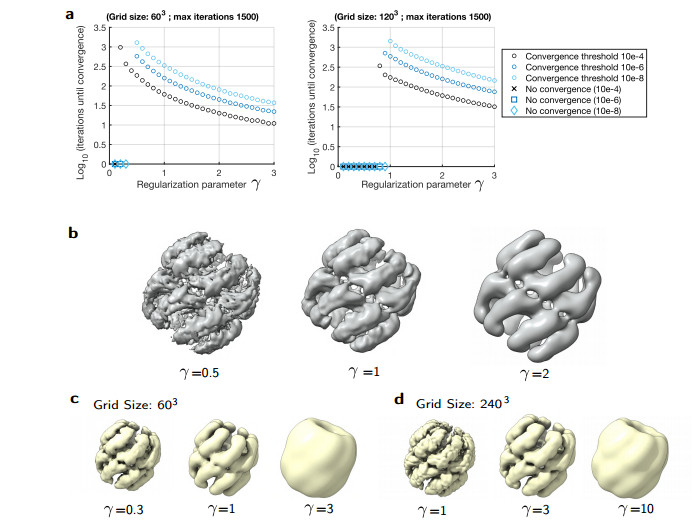
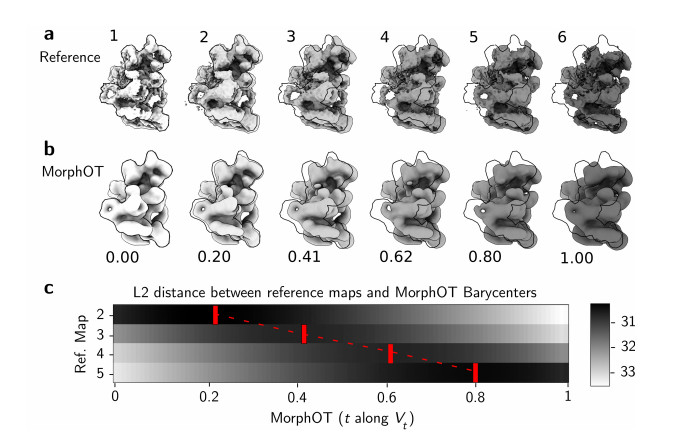
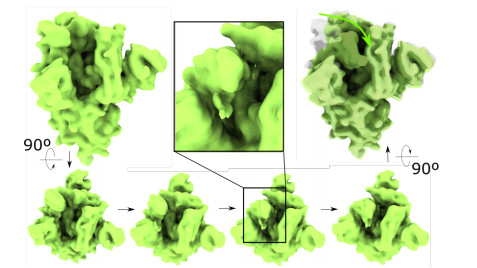
Cryogenic electron microscopy (cryo-EM) has become widely used for the past few years in structural biology, to collect single images of macromolecules "frozen in time". As this technique facilitates the identification of multiple conformational states adopted by the same molecule, a direct product of it is a set of 3D volumes, also called EM maps. To gain more insights on the possible mechanisms that govern transitions between different states, and hence the mode of action of a molecule, we recently introduced a bioinformatic tool that interpolates and generates morphing trajectories joining two given EM maps. This tool is based on recent advances made in optimal transport, that allow efficient evaluation of Wasserstein barycenters of 3D shapes. As the overall performance of the method depends on various key parameters, including the sensitivity of the regularization parameter, we performed various numerical experiments to demonstrate how MorphOT can be applied in different contexts and settings. Finally, we discuss current limitations and further potential connections between other optimal transport theories and the conformational heterogeneity problem inherent with cryo-EM data.
Citation: Arthur Ecoffet, Geoffrey Woollard, Artem Kushner, Frédéric Poitevin, Khanh Dao Duc. Application of transport-based metric for continuous interpolation between cryo-EM density maps[J]. AIMS Mathematics, 2022, 7(1): 986-999. doi: 10.3934/math.2022059
Cryogenic electron microscopy (cryo-EM) has become widely used for the past few years in structural biology, to collect single images of macromolecules "frozen in time". As this technique facilitates the identification of multiple conformational states adopted by the same molecule, a direct product of it is a set of 3D volumes, also called EM maps. To gain more insights on the possible mechanisms that govern transitions between different states, and hence the mode of action of a molecule, we recently introduced a bioinformatic tool that interpolates and generates morphing trajectories joining two given EM maps. This tool is based on recent advances made in optimal transport, that allow efficient evaluation of Wasserstein barycenters of 3D shapes. As the overall performance of the method depends on various key parameters, including the sensitivity of the regularization parameter, we performed various numerical experiments to demonstrate how MorphOT can be applied in different contexts and settings. Finally, we discuss current limitations and further potential connections between other optimal transport theories and the conformational heterogeneity problem inherent with cryo-EM data.

| [1] |
T. Nakane, A. Kotecha, A. Sente, G. McMullan, S. Masiulis, P. MGE Brown, et al., Single-particle cryo-EM at atomic resolution, Nature, 587 (2020), 152–156. doi: 10.1038/s41586-020-2829-0. doi: 10.1038/s41586-020-2829-0

|
| [2] |
D. Lyumkis, Challenges and opportunities in cryo-EM single-particle analysis, J. Biol. Chem., 294 (2019), 5181–5197. doi: 10.1074/jbc.REV118.005602. doi: 10.1074/jbc.REV118.005602
|
| [3] |
F. Poitevin, A. Kushner, X. Li, K. Dao Duc, Structural Heterogeneities of the Ribosome: New Frontiers and Opportunities for Cryo-EM, Molecules, 25 (2020), 4262. doi: 10.3390/molecules25184262. doi: 10.3390/molecules25184262
|
| [4] | J. Frank, Three-dimensional electron microscopy of macromolecular assemblies: visualization of biological molecules in their native state, 2$^{nd}$ edition, Oxford University Press, New York, 2006. |
| [5] |
D. Weiss, M. Levitt, Can morphing methods predict intermediate structures? J. Mol. Biol., 385 (2009), 665–674. doi: 10.1016/j.jmb.2008.10.064. doi: 10.1016/j.jmb.2008.10.064
|
| [6] |
A. Ecoffet, F. Poitevin, K. Dao Duc, MorphOT: Transport-based interpolation between EM maps with UCSF ChimeraX, Bioinformatics, 36 (2020), 5528–5529. doi: 10.1093/bioinformatics/btaa1019. doi: 10.1093/bioinformatics/btaa1019
|
| [7] |
T. Goddard, C. Huang, E. Meng, E. Pettersen, G. Couch, J. Morris, et al., UCSF ChimeraX: Meeting modern challenges in visualization and analysis, Protein Sci., 27 (2018), 14–25. doi: 10.1002/pro.3235. doi: 10.1002/pro.3235
|
| [8] |
J. Solomon, F. De Goes, G. Peyré, M. Cuturi, A. Butscher, A. Nguyen, et al., Convolutional wasserstein distances: Efficient optimal transportation on geometric domains, ACM T. Graphic., 34 (2015), 1–11. doi: 10.1145/2766963. doi: 10.1145/2766963
|
| [9] |
G. Peyré, M. Cuturi, Computational Optimal Transport: With Applications to Data Science, Found. Trends Mach. Learn., 11 (2019), 355–607. doi: 10.1561/2200000073. doi: 10.1561/2200000073
|
| [10] |
R. McCann, A convexity principle for interacting gases, Adv. Math., 128 (1997), 153–179. doi: 10.1006/aima.1997.1634. doi: 10.1006/aima.1997.1634
|
| [11] |
N. Bonneel, M. Van De Panne, S. Paris, W. Heidrich, Displacement interpolation using Lagrangian mass transport, Proceedings of the 2011 SIGGRAPH Asia Conference, 30 (2011), 158. doi: 10.1145/2070781.2024192. doi: 10.1145/2070781.2024192
|
| [12] |
M. Cuturi, Sinkhorn distances: Lightspeed computation of optimal transport, Adv. Neural. Inf. Process. Syst., 26 (2013), 2292–2300. doi: 10.5555/2999792.2999868. doi: 10.5555/2999792.2999868
|
| [13] |
L. M. Bregman, The relaxation method of finding the common point of convex sets and its application to the solution of problems in convex programming, USSR Comput. Math. & Math. Phys., 7 (1967), 200–217. doi: 10.1016/0041-5553(67)90040-7. doi: 10.1016/0041-5553(67)90040-7
|
| [14] | R. M. Glaeser, E. Nogales, W. Chiu, Single-particle Cryo-EM of Biological Macromolecules, IOP Publishing, Bristol, UK, 2021. |
| [15] | R. Flamary, N. Courty, POT Python Optimal Transport library, 2017. Available from: https://pythonot.github.io/. |
| [16] |
P. Virtanen, R. Gommers, T. E. Oliphant, M. Haberland, T. Reddy, D. Cournapeau, et al., SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python, Nat. Methods, 17 (2020), 261–272. doi: 10.1038/s41592-019-0686-2. doi: 10.1038/s41592-019-0686-2
|
| [17] |
J. Zhang, M. L. Baker, G. F. Schröder, N. R. Douglas, S. Reissmann, J. Jakana, et al., Mechanism of folding chamber closure in a group Ⅱ chaperonin, Nature, 463 (2010), 379–383. doi: 10.1038/nature08701. doi: 10.1038/nature08701
|
| [18] |
W. Krebs, M. Gerstein, SURVEY AND SUMMARY: The morph server: a standardized system for analyzing and visualizing macromolecular motions in a database framework, Nucleic Acids Res., 28 (2000), 1665–1675. doi: 10.1093/nar/28.8.1665. doi: 10.1093/nar/28.8.1665
|
| [19] |
G. D. Pintilie, J. Zhang, T. D. Goddard, W. Chiu, D. C. Gossard, Quantitative analysis of cryo-EM density map segmentation by watershed and scale-space filtering, and fitting of structures by alignment to regions, J. Struct. Biol., 170 (2010), 427–438. doi: 10.1016/j.jsb.2010.03.007. doi: 10.1016/j.jsb.2010.03.007
|
| [20] | R. Okuta, Y. Unno, D. Nishino, S. Hido, C. Loomis, CuPy : A NumPy-Compatible Library for NVIDIA GPU Calculations, 31st Confernce on Neural Information Processing Systems, 6 (2017). |
| [21] |
B. Schmitzer, Stabilized Sparse Scaling Algorithms for Entropy Regularized Transport Problems, SIAM J. Sci. Comput., 41 (2019), A1443–A1481. doi: 10.1137/16M1106018. doi: 10.1137/16M1106018
|
| [22] |
E. D. Zhong, T. Bepler, B. Berger, J. H. Davis, CryoDRGN: reconstruction of heterogeneous cryo-EM structures using neural networks, Nat. Methods, 18 (2021), 176–185. doi: 10.1038/s41592-020-01049-4. doi: 10.1038/s41592-020-01049-4
|
| [23] |
D. Haselbach, I. Komarov, D. E. Agafonov, K. Hartmuth, B. Graf, O. Dybkov, et al., Structure and Conformational Dynamics of the Human Spliceosomal Bact Complex, Cell, 172 (2018), 454–464. doi: 10.1016/j.cell.2018.01.010. doi: 10.1016/j.cell.2018.01.010
|
| [24] | E. D. Zhong, Data for "CryoDRGN: Reconstruction of heterogeneous cryo-EM structures using neural networks", Zenodo, 2021. Available from: https://zenodo.org/record/4355284. |
| [25] |
C. O. S. Sorzano, J. M. Carazo, Principal component analysis is limited to low-resolution analysis in cryoEM, Acta Crystallogr. D Struct. Biol., 77 (2021), 835–839. doi: 10.1107/s2059798321002291. doi: 10.1107/s2059798321002291
|
| [26] | A. Punjani, D. Fleet, 3D Flexible Refinement : Structure and Motion of Flexible Proteins from Cryo-EM, preprint, BiorXiv. |
| [27] |
R. Henderson, R. J. Edwards, K. Mansouri, K. Janowska, V. Stalls, S. M. Gobeil, et al., Controlling the SARS-CoV-2 spike glycoprotein conformation, Nat. Struct. Mol. Biol., 27 (2020), 925–933. doi: 10.1038/s41594-020-0479-4. doi: 10.1038/s41594-020-0479-4
|
| [28] |
N. Zelesko, A. Moscovich, J. Kileel, A. Singer, Earthmover-based manifold learning for analyzing molecular conformation spaces, IEEE 17th International Symposium on Biomedical Imaging, (2020), 1715–1719. doi: 10.1109/ISBI45749.2020.9098723. doi: 10.1109/ISBI45749.2020.9098723
|
| [29] | J. Kileel, A. Moscovich, N. Zelesko, A. Singer, Manifold learning with arbitrary norms, preprint, arXiv: 2012.14172. |
| [30] | F. Santambrogio, Optimal Transport for Applied Mathematicians: Calculus of Variations, PDEs, and Modeling, Birkäuser, Springer, New York, 2015. doi: 10.1007/978-3-319-20828-2. |
| [31] |
T. Nakane, D. Kimanius, E. Lindahl, S. Scheres, Characterisation of molecular motions in cryo-EM single-particle data by multi-body refinement in RELION, Elife, 7 (2018), e36861. doi: 10.7554/eLife.36861. doi: 10.7554/eLife.36861
|
| [32] |
L. Chizat, G. Peyré, B. Schmitzer, F. X. Vialard, Scaling algorithms for unbalanced optimal transport problems, Math. Comput., 87 (2018), 2563–2609. doi: 10.1090/mcom/3303. doi: 10.1090/mcom/3303
|
| [33] |
P. Koehl, M. Delarue, H. Orland, Physics approach to the variable-mass optimal-transport problem, Phys. Rev. E, 103 (2021), 012113. doi: 10.1103/PhysRevE.103.012113. doi: 10.1103/PhysRevE.103.012113
|
| [34] |
A. Dupuy, A. Galichon, Personality Traits and the Marriage Market, J. Political Econ., 122 (2014), 1271–1319. doi: 10.1086/677191. doi: 10.1086/677191
|
| [35] | L. Xu, H. Sun, Y. Liu, Learning with Batch-wise Optimal Transport Loss for 3D Shape Recognition, preprint, arXiv: 1903.08923. |
| [36] |
M. Heitz, N. Bonneel, D. Coeurjolly, M. Cuturi, G. Peyré, Ground Metric Learning on Graphs, J. Math. Imaging Vis., 63 (2021), 89–107. doi: 10.1007/s10851-020-00996-z. doi: 10.1007/s10851-020-00996-z
|
| [37] | R. Liu, A. Balsubramani, J. Zou, Learning transport cost from subset correspondence, preprint, arXiv: 1909.13203. |
| [38] | A. Stuart, MT. Wolfram, Inverse optimal transport, preprint, arXiv: 1905.03950. |
| [39] | H. Sun, H. Zhou, H. Zha, X. Ye, Learning Cost Functions for Optimal Transport, preprint, arXiv: 2002.09650. |
Figures(4) / Tables(1)

Arthur Ecoffet, Geoffrey Woollard, Artem Kushner, Frédéric Poitevin, Khanh Dao Duc. Application of transport-based metric for continuous interpolation between cryo-EM density maps[J]. AIMS Mathematics, 2022, 7(1): 986-999. doi: 10.3934/math.2022059







 DownLoad:
DownLoad: