Psychiatric disorders may extremely impair the quality of life with patients and are important reasons of social disability. Several data have shown that psychiatric disorders are associated with an altered composition of gut microbiota. Dietary intake could determine the microbiota, which contribute to produce various metabolites of fermentation such as short chain fatty acids. Some of the metabolites could result in epigenetic alterations leading to the disease susceptibility. Epigenetic dysfunction is in fact implicated in various psychiatric and neurologic disorders. For example, it has been shown that neuroepigenetic dysregulation occurs in psychiatric disorders including schizophrenia. Several studies have demonstrated that the intestinal microbiome may influence the function of central nervous system. Furthermore, it has been proved that the alterations in the gut microbiota-composition might affect in the bidirectional communication between gut and brain. Similarly, evidences demonstrating the association between psychiatric disorders and the gut microbiota have come from preclinical studies. It is clear that an intricate symbiotic relationship might exist between host and microbe, although the practical significance of the gut microbiota has not yet to be determined. In this review, we have summarized the function of gut microbiota in main psychiatric disorders with respect to the mental health. In addition, we would like to discuss the potential mechanisms of the disorders for the practical diagnosis and future treatment by using bioengineering of microbiota and their metabolites.
Citation: Kurumi Taniguchi, Yuka Ikeda, Nozomi Nagase, Ai Tsuji, Yasuko Kitagishi, Satoru Matsuda. Implications of Gut-Brain axis in the pathogenesis of Psychiatric disorders[J]. AIMS Bioengineering, 2021, 8(4): 243-256. doi: 10.3934/bioeng.2021021
Psychiatric disorders may extremely impair the quality of life with patients and are important reasons of social disability. Several data have shown that psychiatric disorders are associated with an altered composition of gut microbiota. Dietary intake could determine the microbiota, which contribute to produce various metabolites of fermentation such as short chain fatty acids. Some of the metabolites could result in epigenetic alterations leading to the disease susceptibility. Epigenetic dysfunction is in fact implicated in various psychiatric and neurologic disorders. For example, it has been shown that neuroepigenetic dysregulation occurs in psychiatric disorders including schizophrenia. Several studies have demonstrated that the intestinal microbiome may influence the function of central nervous system. Furthermore, it has been proved that the alterations in the gut microbiota-composition might affect in the bidirectional communication between gut and brain. Similarly, evidences demonstrating the association between psychiatric disorders and the gut microbiota have come from preclinical studies. It is clear that an intricate symbiotic relationship might exist between host and microbe, although the practical significance of the gut microbiota has not yet to be determined. In this review, we have summarized the function of gut microbiota in main psychiatric disorders with respect to the mental health. In addition, we would like to discuss the potential mechanisms of the disorders for the practical diagnosis and future treatment by using bioengineering of microbiota and their metabolites.
Alzheimer's disease
attention deficit hyperactivity disorder
autism spectrum disorder
blood brain barrier
central nervous system
deoxyribonucleic acid
gamma amino butyric acid
gastrointestinal
G protein-coupled receptor
histone deacetylases
lipopolysaccharide
reactive oxygen species
superoxide dismutase

| [1] |
Mathee K, Cickovski T, Deoraj A, et al. (2020) The gut microbiome and neuropsychiatric disorders: implications for attention deficit hyperactivity disorder (ADHD). J Med Microbiol 69: 14-24. doi: 10.1099/jmm.0.001112

|
| [2] |
Vigo D, Thornicroft G, Atun R (2016) Estimating the true global burden of mental illness. Lancet Psychiatry 3: 171-178. doi: 10.1016/S2215-0366(15)00505-2
|
| [3] |
Os J, Kenis G, Rutten BP (2010) The environment and schizophrenia. Nature 468: 203-212. doi: 10.1038/nature09563
|
| [4] |
Köhler-Forsberg O, Petersen L, Gasse C, et al. (2019) A nationwide study in denmark of the association between treated infections and the subsequent risk of treated mental disorders in children and adolescents. JAMA Psychiatry 76: 271-279. doi: 10.1001/jamapsychiatry.2018.3428
|
| [5] |
Goldsmith DR, Rapaport MH, Miller BJ (2016) A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Mol Psychiatry 21: 1696-1709. doi: 10.1038/mp.2016.3
|
| [6] |
Krefft M, Frydecka D, Zalsman G, et al. (2021) A pro-inflammatory phenotype is associated with behavioural traits in children with Prader-Willi syndrome. Eur Child Adolesc Psychiatry 30: 899-908. doi: 10.1007/s00787-020-01568-7
|
| [7] |
Shahbazi R, Yasavoli-Sharahi H, Alsadi N, et al. (2020) Matar, C. Probiotics in treatment of viral respiratory infections and neuroinflammatory disorders. Molecules 25: 4891. doi: 10.3390/molecules25214891
|
| [8] |
Barona M, Brown M, Clark C, et al. (2019) White matter alterations in anorexia nervosa: Evidence from a voxel-based meta-analysis. Neurosci Biobehav Rev 100: 285-295. doi: 10.1016/j.neubiorev.2019.03.002
|
| [9] |
Bastiaanssen TFS, Cryan JF (2021) The microbiota-gut-brain axis in mental health and medication response: parsing directionality and causality. Int J Neuropsychopharmacol 24: 216-220. doi: 10.1093/ijnp/pyaa088
|
| [10] |
Martins LB, Braga Tibães JR, Sanches M, et al. (2021) Nutrition-based interventions for mood disorders. Expert Rev Neurother 21: 303-315. doi: 10.1080/14737175.2021.1881482
|
| [11] |
Cerdó T, Diéguez E, Campoy C (2020) Impact of gut microbiota on neurogenesis and neurological diseases during infancy. Curr Opin Pharmacol 50: 33-37. doi: 10.1016/j.coph.2019.11.006
|
| [12] |
Gubert C, Kong G, Renoir T, et al. (2020) Exercise, diet and stress as modulators of gut microbiota: Implications for neurodegenerative diseases. Neurobiol Dis 134: 104621. doi: 10.1016/j.nbd.2019.104621
|
| [13] |
Berding K, Vlckova K, Marx W, et al. Diet and the microbiota-gut-brain axis: sowing the seeds of good mental health (2021) . doi: 10.1093/advances/nmaa181
|
| [14] |
Rea K, Dinan TG, Cryan JF (2020) Gut microbiota: a perspective for psychiatrists. Neuropsychobiology 79: 50-62. doi: 10.1159/000504495
|
| [15] |
Zinchenko E, Navolokin N, Shirokov A, et al. (2019) Pilot study of transcranial photobiomodulation of lymphatic clearance of beta-amyloid from the mouse brain: breakthrough strategies for non-pharmacologic therapy of Alzheimer's disease. Biomed Opt Express 10: 4003-4017. doi: 10.1364/BOE.10.004003
|
| [16] |
Mishra SP, Shukla SK, Pandey BL (2014) A preliminary evaluation of comparative effectiveness of riluzole in therapeutic regimen for irritable bowel syndrome. Asian Pac J Trop Biomed 4: S335-S340. doi: 10.12980/APJTB.4.2014C205
|
| [17] |
Barberio B, Zamani M, Black CJ, et al. (2021) Prevalence of symptoms of anxiety and depression in patients with inflammatory bowel disease: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol 6: 359-370. doi: 10.1016/S2468-1253(21)00014-5
|
| [18] |
Dinan TG, Cryan JF (2017) Gut instincts: microbiota as a key regulator of brain development, ageing and neurodegeneration. J Physiol 595: 489-503. doi: 10.1113/JP273106
|
| [19] |
Wang J, Gu X, Yang J, et al. (2019) Gut microbiota dysbiosis and increased plasma LPS and TMAO levels in patients with preeclampsia. Front Cell Infect Microbiol 9: 409. doi: 10.3389/fcimb.2019.00409
|
| [20] |
Stasi C, Caserta A, Nisita C, et al. (2019) The complex interplay between gastrointestinal and psychiatric symptoms in irritable bowel syndrome: A longitudinal assessment. J Gastroenterol Hepatol 34: 713-719. doi: 10.1111/jgh.14375
|
| [21] |
Kelly JR, Clarke G, Cryan JF, et al. (2016) Brain-gut-microbiota axis: challenges for translation in psychiatry. Ann Epidemiol 26: 366-372. doi: 10.1016/j.annepidem.2016.02.008
|
| [22] |
Winter G, Hart RA, Charlesworth RPG, et al. (2018) Gut microbiome and depression: what we know and what we need to know. Rev Neurosci 29: 629-643. doi: 10.1515/revneuro-2017-0072
|
| [23] |
Zheng P, Zeng B, Zhou C, et al. (2016) Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host's metabolism. Mol Psychiatry 21: 786-796. doi: 10.1038/mp.2016.44
|
| [24] |
Yang C, Qu Y, Fujita Y, et al. (2017) Possible role of the gut microbiota-brain axis in the antidepressant effects of (R)-ketamine1 in a social defeat stress model. Transl Psychiatry 7: 1294. doi: 10.1038/s41398-017-0031-4
|
| [25] |
Lyte M, Daniels KM, Schmitz-Esser S (2019) Fluoxetine-induced alteration of murine gut microbial community structure: evidence for a microbial endocrinology-based mechanism of action responsible for fluoxetine-induced side effects. Peer J 7: e6199. doi: 10.7717/peerj.6199
|
| [26] |
Saini A, Ardine M, Berruti A (2008) Major depressive disorder. N Engl J Med 358: 1868-1869. doi: 10.1056/NEJMc080186
|
| [27] |
Arneth BM (2018) Gut-brain axis biochemical signalling from the gastrointestinal tract to the central nervous system: gut dysbiosis and altered brain function. Postgrad Med J 94: 446-452. doi: 10.1136/postgradmedj-2017-135424
|
| [28] |
Borre YE, O'Keeffe GW, Clarke G, et al. (2014) Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med 20: 509-518. doi: 10.1016/j.molmed.2014.05.002
|
| [29] |
Barbara G, Stanghellini V, Brandi G, et al. (2005) Interactions between commensal bacteria and gut sensorimotor function in health and disease. Am J Gastroenterol 100: 2560-2568. doi: 10.1111/j.1572-0241.2005.00230.x
|
| [30] |
Strandwitz P (2018) Neurotransmitter modulation by the gut microbiota. Brain Res 1693: 128-133. doi: 10.1016/j.brainres.2018.03.015
|
| [31] |
Sivamaruthi BS, Suganthy N, Kesika P, et al. (2020) The role of microbiome, dietary supplements, and probiotics in autism spectrum disorder. Int J Environ Res Public Health 17: 2647. doi: 10.3390/ijerph17082647
|
| [32] |
Song Q, Wang Y, Huang L, et al. (2021) Review of the relationships among polysaccharides, gut microbiota, and human health. Food Res Int 140: 109858. doi: 10.1016/j.foodres.2020.109858
|
| [33] |
Salami M (2021) Interplay of good bacteria and central nervous system: cognitive aspects and mechanistic considerations. Front Neurosci 15: 613120. doi: 10.3389/fnins.2021.613120
|
| [34] |
Niederseer D, Wernly B, Aigner E, et al. (2021) NAFLD and cardiovascular diseases: epidemiological, mechanistic and therapeutic considerations. J Clin Med 10: 467. doi: 10.3390/jcm10030467
|
| [35] |
Burgueño JF, Fritsch J, Santander AM, et al. (2019) Intestinal epithelial cells respond to chronic inflammation and dysbiosis by synthesizing H2O2. Front Physiol 10: 1484. doi: 10.3389/fphys.2019.01484
|
| [36] |
Belizário JE, Faintuch J, Garay-Malpartida M (2018) Gut microbiome dysbiosis and immunometabolism: New frontiers for treatment of metabolic diseases. Mediators Inflamm 2018: 2037838. doi: 10.1155/2018/2037838
|
| [37] | Andreyev AY, Kushnareva YE, Starkova NN, et al. (2020) Metabolic ROS signaling: to immunity and beyond. Biochemistry 85: 1650-1667. |
| [38] | Saint-Georges-Chaumet Y, Attaf D, Pelletier E, et al. (2015) Targeting microbiota-mitochondria inter-talk: Microbiota control mitochondria metabolism. Cell Mol Biol 61: 121-124. |
| [39] |
Li M, Gu MM, Lang Y, et al. (2019) The vanillin derivative VND3207 protects intestine against radiation injury by modulating p53/NOXA signaling pathway and restoring the balance of gut microbiota. Free Radi Biol Med 145: 223-236. doi: 10.1016/j.freeradbiomed.2019.09.035
|
| [40] |
Wang W, Im J, Kim S, et al. (2020) ROS-induced SIRT2 upregulation contributes to cisplatin sensitivity in ovarian cancer. Antioxidants 9: 1137. doi: 10.3390/antiox9111137
|
| [41] |
Muralitharan RR, Marques FZ (2021) Diet-related gut microbial metabolites and sensing in hypertension. J Hum Hypertens 35: 162-169. doi: 10.1038/s41371-020-0388-3
|
| [42] |
Zhou Y, Jiang Q, Zhao S, et al. (2019) Impact of buckwheat fermented milk combined with high-fat diet on rats' gut microbiota and short-chain fatty acids. J Food Sci 84: 3833-3842. doi: 10.1111/1750-3841.14958
|
| [43] |
Skonieczna-Żydecka K, Grochans E, Maciejewska D, et al. (2018) Short chain fatty acids profile is changed in Polish depressive women. Nutrients 10: 1939. doi: 10.3390/nu10121939
|
| [44] |
Hu L, Zhu S, Peng X, et al. (2020) High salt elicits brain inflammation and cognitive dysfunction, accompanied by alternations in the gut microbiota and decreased SCFA production. J Alzheimers Dis 77: 629-640. doi: 10.3233/JAD-200035
|
| [45] |
Luceri C, Femia AP, Fazi M, et al. (2016) Effect of butyrate enemas on gene expression profiles and endoscopic/histopathological scores of diverted colorectal mucosa: A randomized trial. Dig Liver Dis 48: 27-33. doi: 10.1016/j.dld.2015.09.005
|
| [46] | Häselbarth L, Ouwens DM, Teichweyde N, et al. (2016) The small chain fatty acid butyrate antagonizes the TCR-stimulation-induced metabolic shift in murine epidermal gamma delta T cells. EXCLI J 19: 334-350. |
| [47] |
Haase S, Haghikia A, Wilck N, et al. (2018) Impacts of microbiome metabolites on immune regulation and autoimmunity. Immunology 154: 230-238. doi: 10.1111/imm.12933
|
| [48] |
Macfarlane GT, Macfarlane S (2012) Bacteria, colonic fermentation, and gastrointestinal health. J AOAC Int 95: 50-60. doi: 10.5740/jaoacint.SGE_Macfarlane
|
| [49] |
Ray S, Das S, Panda PK, et al. (2018) Identification of a new alanine racemase in Salmonella Enteritidis and its contribution to pathogenesis. Gut Pathog 10: 30. doi: 10.1186/s13099-018-0257-6
|
| [50] |
Gilmore MS, Skaugen M, Nes I (1996) Enterococcus faecalis cytolysin and lactocin S of Lactobacillus sake. Antonie Van Leeuwenhoek 69: 129-138. doi: 10.1007/BF00399418
|
| [51] |
Pidgeon SE, Fura JM, Leon W, et al. (2015) Metabolic profiling of bacteria by unnatural C-terminated d-Amino Acids. Angew Chem 127: 6256-6260. doi: 10.1002/ange.201409927
|
| [52] |
Sasabe J, Miyoshi Y, Rakoff-Nahoum S, et al. (2016) Interplay between microbial d-amino acids and host d-amino acid oxidase modifies murine mucosal defence and gut microbiota. Nat Microbiol 1: 16125. doi: 10.1038/nmicrobiol.2016.125
|
| [53] |
Nagano T, Yamao S, Terachi A, et al. (2019) d-amino acid oxidase promotes cellular senescence via the production of reactive oxygen species. Life Sci Alliance 2: e201800045. doi: 10.26508/lsa.201800045
|
| [54] |
Zhong C, Zhu N, Zhu Y, et al. (2020) Antimicrobial peptides conjugated with fatty acids on the side chain of D-amino acid promises antimicrobial potency against multidrug-resistant bacteria. Eur J Pharm Sci 141: 105123. doi: 10.1016/j.ejps.2019.105123
|
| [55] |
Arai T, Ashraful Hoque M, Nishino N, et al. (2013) Cyclic tetrapeptides with -SS- bridging between amino acid side chains for potent histone deacetylases' inhibition. Amino Acids 45: 835-843. doi: 10.1007/s00726-013-1527-8
|
| [56] |
Hergueta T, Weiller E (2013) Evaluating depressive symptoms in hypomanic and manic episodes using a structured diagnostic tool: validation of a new Mini International Neuropsychiatric Interview (M.I.N.I.) module for the DSM-5 ‘With Mixed Features’ specifier. Int J Bipolar Disord 1: 21. doi: 10.1186/2194-7511-1-21
|
| [57] |
Ryu JC, Zimmer ER, Rosa-Neto P, et al. (2019) Consequences of metabolic disruption in Alzheimer's disease pathology. Neurotherapeutics 16: 600-610. doi: 10.1007/s13311-019-00755-y
|
| [58] |
Laversenne V, Nazeeruddin S, Källstig EC, et al. (2020) Anti-Aβ antibodies bound to neuritic plaques enhance microglia activity and mitigate tau pathology. Acta Neuropathol Commun 8: 198. doi: 10.1186/s40478-020-01069-3
|
| [59] |
Guo LX, Tong Y, Wang J, et al. (2020) Determination and comparison of short-chain fatty acids in serum and colon content samples: Alzheimer's disease rat as a case study. Molecules 25: 5739. doi: 10.3390/molecules25235739
|
| [60] |
Liu H, Zhang JJ, Li X, et al. (2015) Post-occlusion administration of sodium butyrate attenuates cognitive impairment in a rat model of chronic cerebral hypoperfusion. Pharmacol Biochem Behav 135: 53-59. doi: 10.1016/j.pbb.2015.05.012
|
| [61] |
Infante R, Scaglione C, Incensi A, et al. (2020) Longitudinal skin biopsy study of phosphorylated alpha-synuclein in a patient with Parkinson disease and orthostatic hypotension. J Neuropathol Exp Neurol 79: 813-816. doi: 10.1093/jnen/nlaa048
|
| [62] |
Mannal N, Kleiner K, Fauler M, et al. (2021) Multi-electrode array analysis identifies complex dopamine responses and glucose sensing properties of substantia nigra neurons in mouse brain slices. Front Synaptic Neurosci 13: 635050. doi: 10.3389/fnsyn.2021.635050
|
| [63] |
Unger MM, Spiegel J, Dillmann KU, et al. (2016) Short chain fatty acids and gut microbiota differ between patients with Parkinson's disease and age-matched controls. Parkinsonism Relat Disord 32: 66-72. doi: 10.1016/j.parkreldis.2016.08.019
|
| [64] |
Ostendorf F, Metzdorf J, Gold R, et al. (2020) Propionic acid and fasudil as treatment against rotenone toxicity in an in vitro model of Parkinson's disease. Molecules 25: 2502. doi: 10.3390/molecules25112502
|
| [65] |
Ostendorf F, Metzdorf J, Gold R, et al. (2020) Differential effects of Δ9-tetrahydrocannabinol dosing on correlates of schizophrenia in the sub-chronic PCP rat model. PLoS One 15: e0230238. doi: 10.1371/journal.pone.0230238
|
| [66] |
Gubert C, Kong G, Uzungil V, et al. (2020) Microbiome profiling reveals gut dysbiosis in the metabotropic glutamate receptor 5 knockout mouse model of schizophrenia. Front Cell Dev Biol 8: 582320. doi: 10.3389/fcell.2020.582320
|
| [67] |
Hu M, Zheng P, Xie Y, et al. (2018) Propionate protects haloperidol-induced neurite lesions mediated by neuropeptide Y. Front Neurosci 12: 743. doi: 10.3389/fnins.2018.00743
|
| [68] |
Girardi Paskulin LM, Kottwitz Bierhals CCB, Oliveira Dos Santos N, et al. (2017) Depressive symptoms of the elderly people and caregiver's burden in home care. Invest Educ Enferm 35: 210-220. doi: 10.17533/udea.iee.v35n2a10
|
| [69] |
Wu M, Tian T, Mao Q, et al. (2020) Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl Psychiatry 10: 350. doi: 10.1038/s41398-020-01038-3
|
| [70] |
Mendonça IP, Duarte-Silva E, Chaves-Filho AJM, et al. (2020) Neurobiological findings underlying depressive behavior in Parkinson's disease: A review. Int Immunopharmacol 83: 106434. doi: 10.1016/j.intimp.2020.106434
|
| [71] |
Maigoro AY, Lee S (2021) Gut microbiome-based analysis of lipid a biosynthesis in individuals with autism spectrum disorder: an in ailico valuation. Nutrients 13: 688. doi: 10.3390/nu13020688
|
| [72] |
Rose DR, Yang H, Serena G, et al. (2018) Differential immune responses and microbiota profiles in children with autism spectrum disorders and co-morbid gastrointestinal symptoms. Brain Behav Immun 70: 354-368. doi: 10.1016/j.bbi.2018.03.025
|
| [73] |
Huo W, Qi P, Cui L, et al. (2020) Polysaccharide from wild morels alters the spatial structure of gut microbiota and the production of short-chain fatty acids in mice. Biosci Microbiota Food Health 39: 219-226. doi: 10.12938/bmfh.2020-018
|
| [74] |
Dickerson FB, Stallings C, Origoni A, et al. Effect of probiotic supplementation on schizophrenia symptoms and association with gastrointestinal functioning: a randomized, placebo-controlled trial (2014) . doi: 10.4088/PCC.13m01579
|
| [75] |
Varela RB, Valvassori SS, Lopes-Borges J, et al. (2015) Sodium butyrate and mood stabilizers block ouabain-induced hyperlocomotion and increase BDNF, NGF and GDNF levels in brain of Wistar rats. J Psychiatr Res 61: 114-121. doi: 10.1016/j.jpsychires.2014.11.003
|
| [76] |
Yamawaki Y, Yoshioka N, Nozaki K, et al. (2018) Sodium butyrate abolishes lipopolysaccharide-induced depression-like behaviors and hippocampal microglial activation in mice. Brain Res 1680: 13-38. doi: 10.1016/j.brainres.2017.12.004
|
| [77] |
Lee J, Venna VR, Durgan DJ, et al. (2020) Young versus aged microbiota transplants to germ-free mice: increased short-chain fatty acids and improved cognitive performance. Gut Microbes 12: 1814107. doi: 10.1080/19490976.2020.1814107
|
| [78] |
D'Mello SR (2019) Regulation of central nervous system development by class I histone deacetylases. Dev Neurosci 41: 149-165. doi: 10.1159/000505535
|
| [79] |
Sinn DI, Kim SJ, Chu K, et al. (2007) Valproic acid-mediated neuroprotection in intracerebral hemorrhage via histone deacetylase inhibition and transcriptional activation. Neurobiol Dis 26: 464-472. doi: 10.1016/j.nbd.2007.02.006
|
| [80] |
Nestler EJ, Peña CJ, Kundakovic M, et al. (2016) Epigenetic basis of mental illness. Neuroscientist 22: 447-463. doi: 10.1177/1073858415608147
|
| [81] |
Kwon B, Houpt TA (2010) Phospho-acetylation of histone H3 in the amygdala after acute lithium chloride. Brain Res 1333: 36-47. doi: 10.1016/j.brainres.2010.03.068
|
| [82] |
Xiang B, Liu K, Yu M, et al. (2018) Systematic genetic analyses of genome-wide association study data reveal an association between the key nucleosome remodeling and deacetylase complex and bipolar disorder development. Bipolar Disord 20: 370-380. doi: 10.1111/bdi.12580
|
| [83] |
Arent CO, Valvassori SS, Fries GR, et al. (2011) Neuroanatomical profile of antimaniac effects of histone deacetylases inhibitors. Mol Neurobiol 43: 207-214. doi: 10.1007/s12035-011-8178-0
|
| [84] |
Guidotti A, Auta J, Chen Y, et al. (2011) Epigenetic GABAergic targets in schizophrenia and bipolar disorder. Neuropharmacology 60: 1007-1016. doi: 10.1016/j.neuropharm.2010.10.021
|
| [85] |
Kramer JM (2013) Epigenetic regulation of memory: implications in human cognitive disorders. Biomol Concepts 4: 1-12. doi: 10.1515/bmc-2012-0026
|
| [86] |
Martínez-Pacheco H, Picazo O, López-Torres A, et al. (2020) Biochemical and Behavioral Characterization of IN14, a New Inhibitor of HDACs with Antidepressant-Like Properties. Biomolecules 10: 299. doi: 10.3390/biom10020299
|
| [87] |
Cuadrado-Tejedor M, Pérez-González M, García-Muñoz C, et al. (2019) Taking advantage of the selectivity of histone deacetylases and phosphodiesterase inhibitors to design better therapeutic strategies to treat Alzheimer's disease. Front Aging Neurosci 11: 149. doi: 10.3389/fnagi.2019.00149
|
| [88] |
Mazzocchi M, Collins LM, Sullivan AM, et al. (2020) The class II histone deacetylases as therapeutic targets for Parkinson's disease. Neuronal Signal 4: NS20200001. doi: 10.1042/NS20200001
|
| [89] |
Marsh JL, Lukacsovich T, Thompson LM (2009) Animal models of polyglutamine diseases and therapeutic approaches. J Biol Chem 284: 7431-7435. doi: 10.1074/jbc.R800065200
|
| [90] |
Frost G, Sleeth ML, Sahuri-Arisoylu M, et al. (2014) The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun 5: 3611. doi: 10.1038/ncomms4611
|
| [91] |
Braniste V, Al-Asmakh M, Kowal C, et al. (2014) The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med 6: 263ra158. doi: 10.1126/scitranslmed.3009759
|
| [92] |
Zhou Z, Xu N, Matei N, et al. (2021) Sodium butyrate attenuated neuronal apoptosis via GPR41/Gbetagamma/PI3K/Akt pathway after MCAO in rats. J Cereb Blood Flow Metab 41: 267-281. doi: 10.1177/0271678X20910533
|
| [93] |
Simonini MV, Camargo LM, Dong E, et al. (2006) The benzamide MS-275 is a potent, long-lasting brain region-selective inhibitor of histone deacetylases. Proc Natl Acad Sci USA 103: 1587-1592. doi: 10.1073/pnas.0510341103
|
| [94] |
Tseng CJ, Gilbert TM, Catanese MC, et al. (2020) In vivo human brain expression of histone deacetylases in bipolar disorder. Transl Psychiatry 10: 224. doi: 10.1038/s41398-020-00911-5
|
| [95] |
Kennedy H, Montreuil TC (2021) The late positive potential as a reliable neural marker of cognitive reappraisal in children and youth: a brief review of the research literature. Front Psychol 11: 608522. doi: 10.3389/fpsyg.2020.608522
|
| [96] |
Gilbert TM, Zürcher NR, Wu CJ, et al. (2019) PET neuroimaging reveals histone deacetylase dysregulation in schizophrenia. J Clin Invest 129: 364-372. doi: 10.1172/JCI123743
|
| [97] |
Wang C, Schroeder FA, Hooker JM (2014) Visualizing epigenetics: current advances and advantages in HDAC PET imaging techniques. Neuroscience 264: 186-197. doi: 10.1016/j.neuroscience.2013.09.018
|
| [98] |
Seo YJ, Muench L, Reid A, et al. (2013) Radionuclide labeling and evaluation of candidate radioligands for PET imaging of histone deacetylase in the brain. Bioorg Med Chem Lett 23: 6700-6705. doi: 10.1016/j.bmcl.2013.10.038
|
| [99] |
Gilbert TM, Zürcher NR, Catanese MC, et al. (2019) Neuroepigenetic signatures of age and sex in the living human brain. Nat Commun 10: 2945. doi: 10.1038/s41467-019-11031-0
|
| [100] |
Omori K, Miyakawa H, Watanabe A, et al. (2021) The combined effects of magnesium oxide and inulin on intestinal microbiota and cecal short-chain fatty acids. Nutrients 13: 152. doi: 10.3390/nu13010152
|
| [101] |
Minami NS, Sousa RS, Oliveira FLC, et al. (2020) Subacute ruminal acidosis in zebu cattle: clinical and behavioral aspects. Animals 11: 21. doi: 10.3390/ani11010021
|
| [102] |
Takewaki D, Suda W, Sato W, et al. (2020) Alterations of the gut ecological and functional microenvironment in different stages of multiple sclerosis. Proc Natl Acad Sci USA 117: 22402-22412. doi: 10.1073/pnas.2011703117
|
| [103] |
Borgo F, Riva A, Benetti A, et al. (2017) Microbiota in anorexia nervosa: The triangle between bacterial species, metabolites and psychological tests. PLoS One 12: e0179739. doi: 10.1371/journal.pone.0179739
|
| [104] |
Geirnaert A, Calatayud M, Grootaert C, et al. (2017) Butyrate-producing bacteria supplemented in vitro to Crohn's disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci Rep 7: 11450. doi: 10.1038/s41598-017-11734-8
|
| [105] | Kilinçarslan S, Evrensel A (2020) The effect of fecal microbiota transplantation on psychiatric symptoms among patients with inflammatory bowel disease: an experimental study. Actas Esp Psiquiatr 48: 1-7. |
| [106] | Tjellström B, Högberg L, Stenhammar L, et al. (2013) Faecal short-chain fatty acid pattern in childhood coeliac disease is normalised after more than one year's gluten-free diet. Microb Ecol Health Dis 24: 20905. |
| [107] | Lerner A, Freire de Carvalho J, Kotrova A, et al. (2021) Gluten-free diet can ameliorate the symptoms of non-celiac autoimmune diseases . |
| [108] |
Mohan M, Okeoma CM, Sestak K (2020) Dietary gluten and neurodegeneration: a case for preclinical studies. Int J Mol Sci 21: 5407. doi: 10.3390/ijms21155407
|
| [109] |
Bruce-Keller AJ, Salbaum JM, Luo M, et al. (2015) Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol Psychiatry 77: 607-615. doi: 10.1016/j.biopsych.2014.07.012
|
| [110] |
Simeonova D, Ivanovska M, Murdjeva M, et al. (2018) Recognizing the leaky gut as a trans-diagnostic target for neuroimmune disorders using clinical chemistry and molecular immunology assays. Curr Top Med Chem 18: 1641-1655. doi: 10.2174/1568026618666181115100610
|
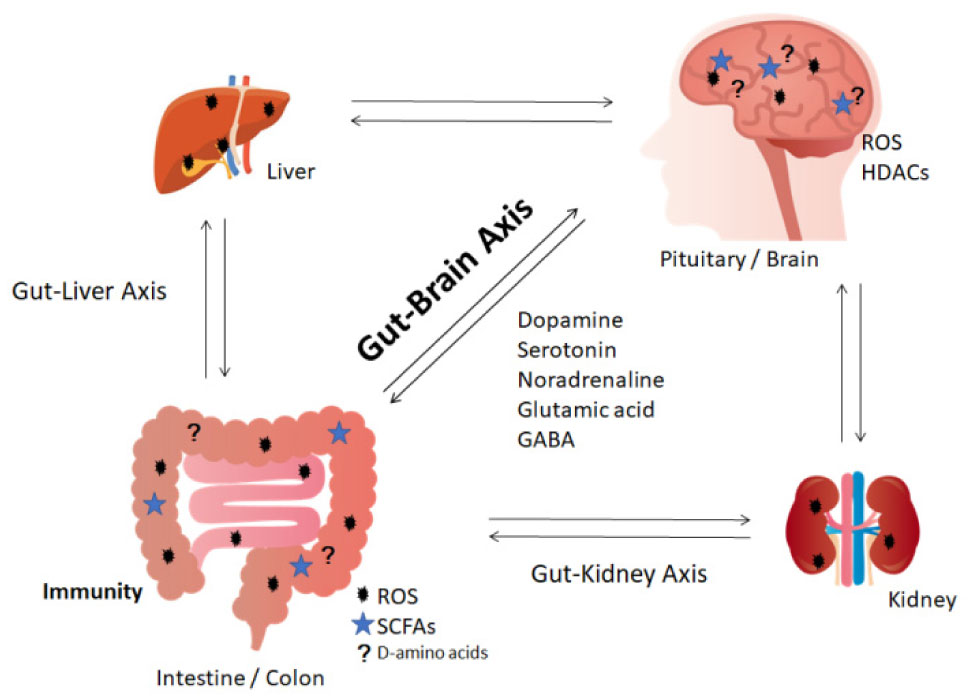
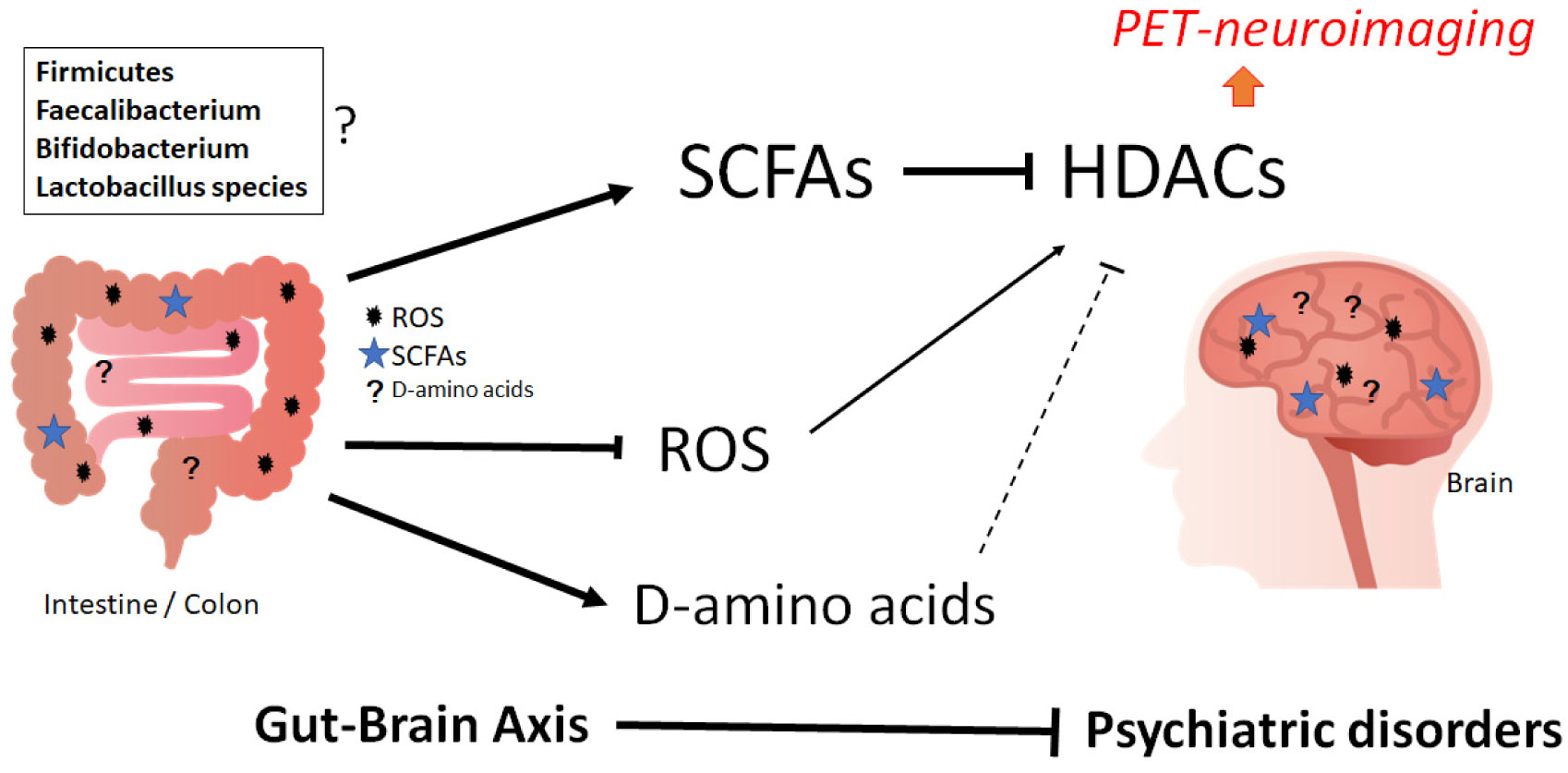
Figures(2)

Kurumi Taniguchi, Yuka Ikeda, Nozomi Nagase, Ai Tsuji, Yasuko Kitagishi, Satoru Matsuda. Implications of Gut-Brain axis in the pathogenesis of Psychiatric disorders[J]. AIMS Bioengineering, 2021, 8(4): 243-256. doi: 10.3934/bioeng.2021021







 DownLoad:
DownLoad: