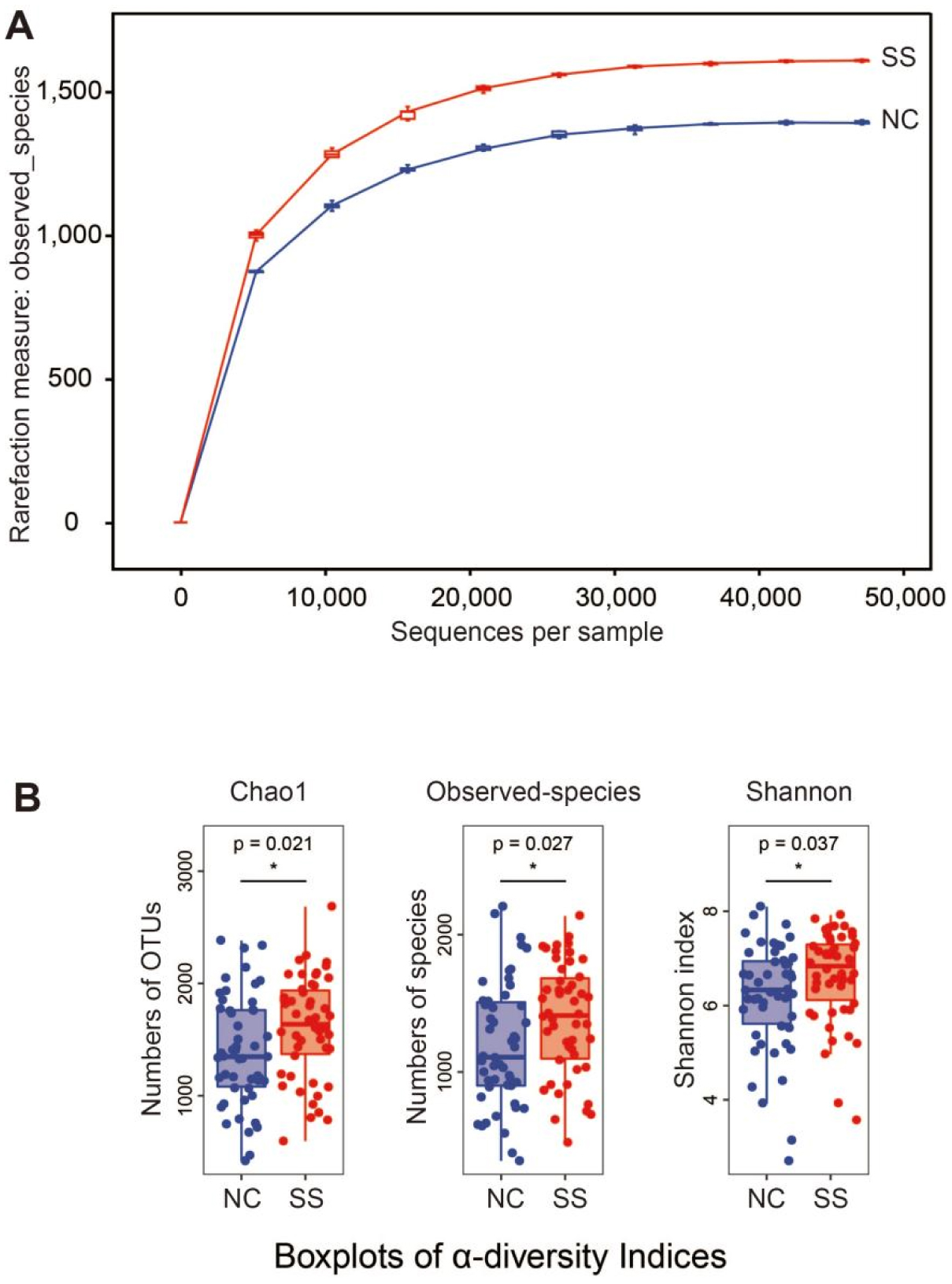
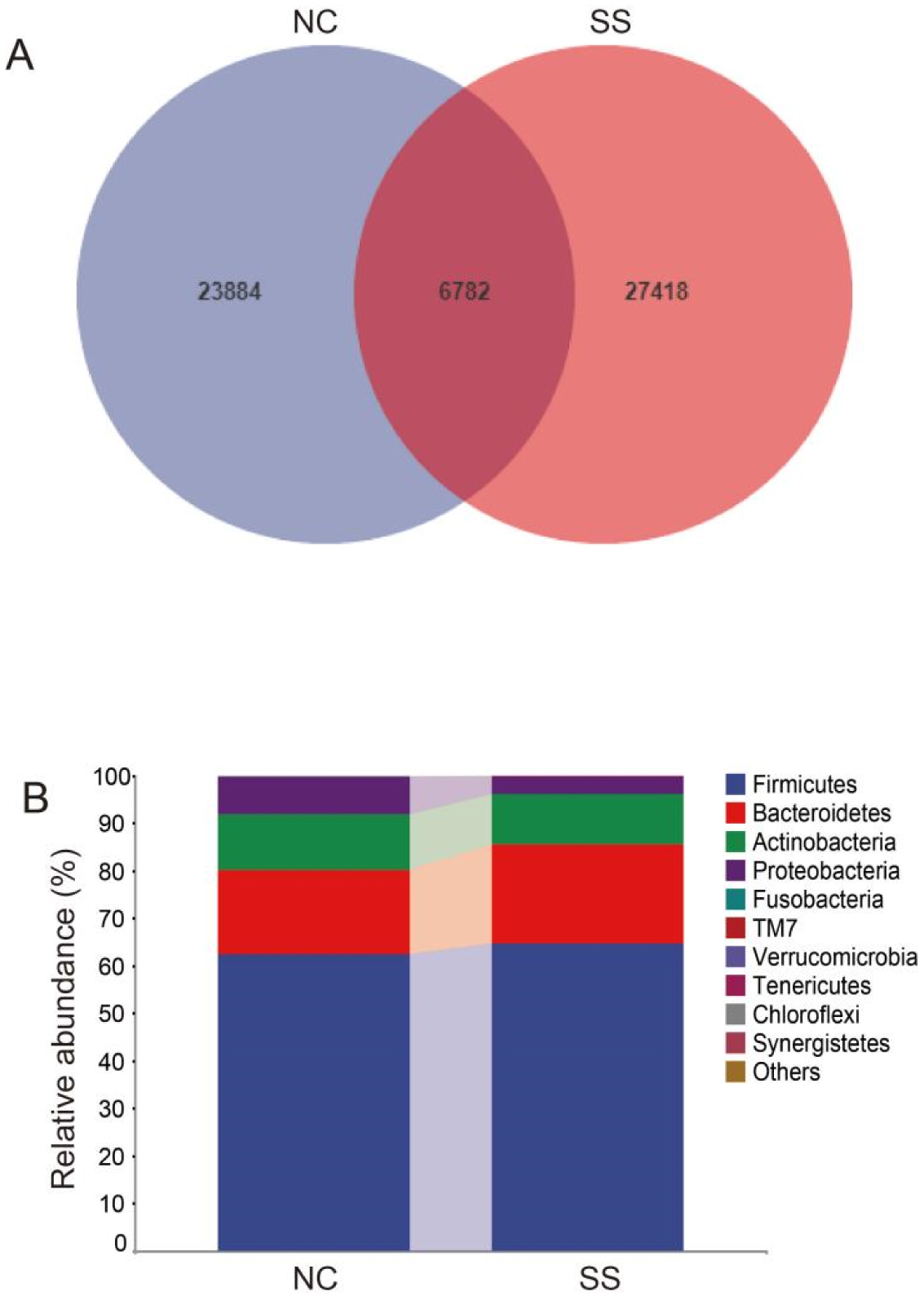
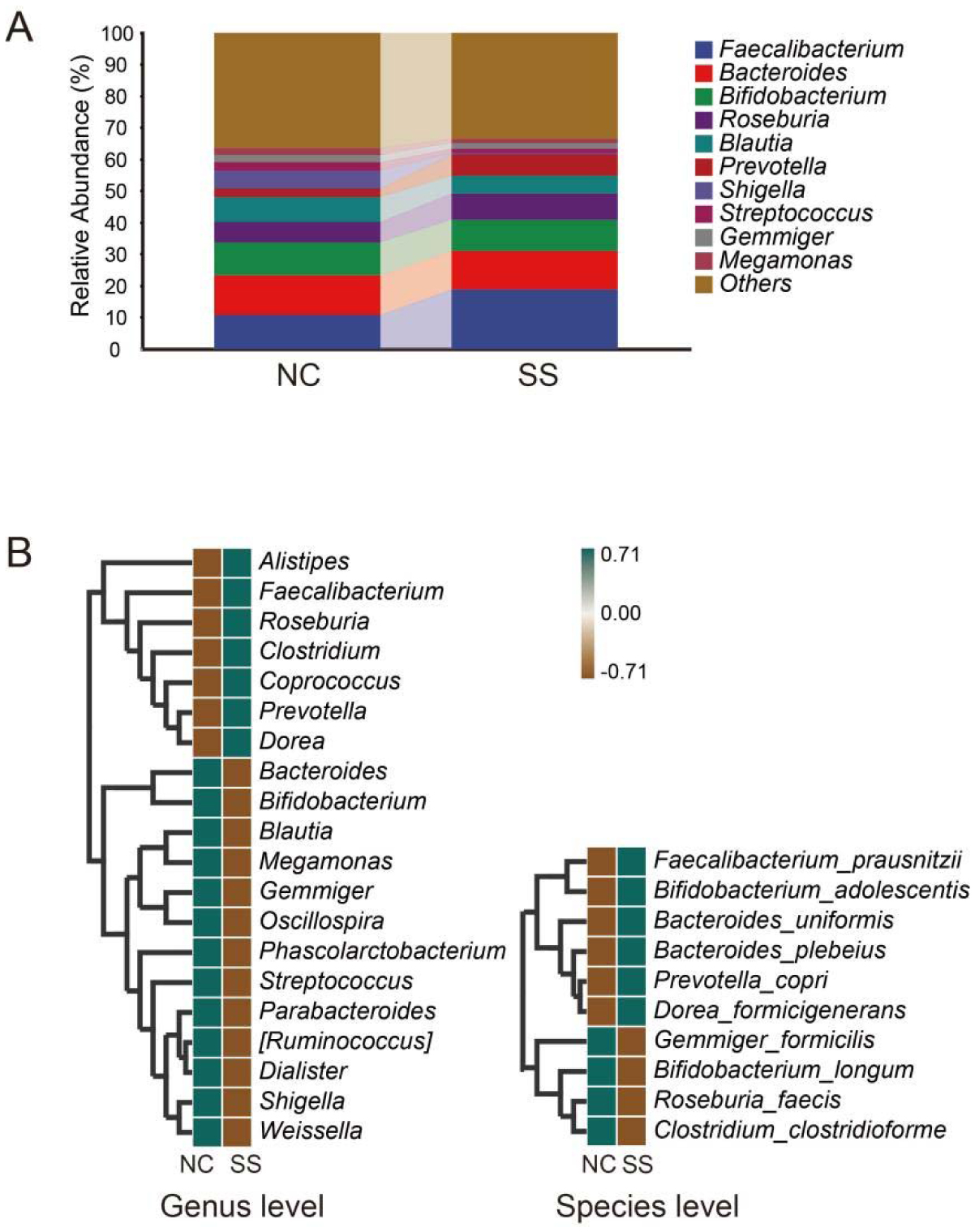
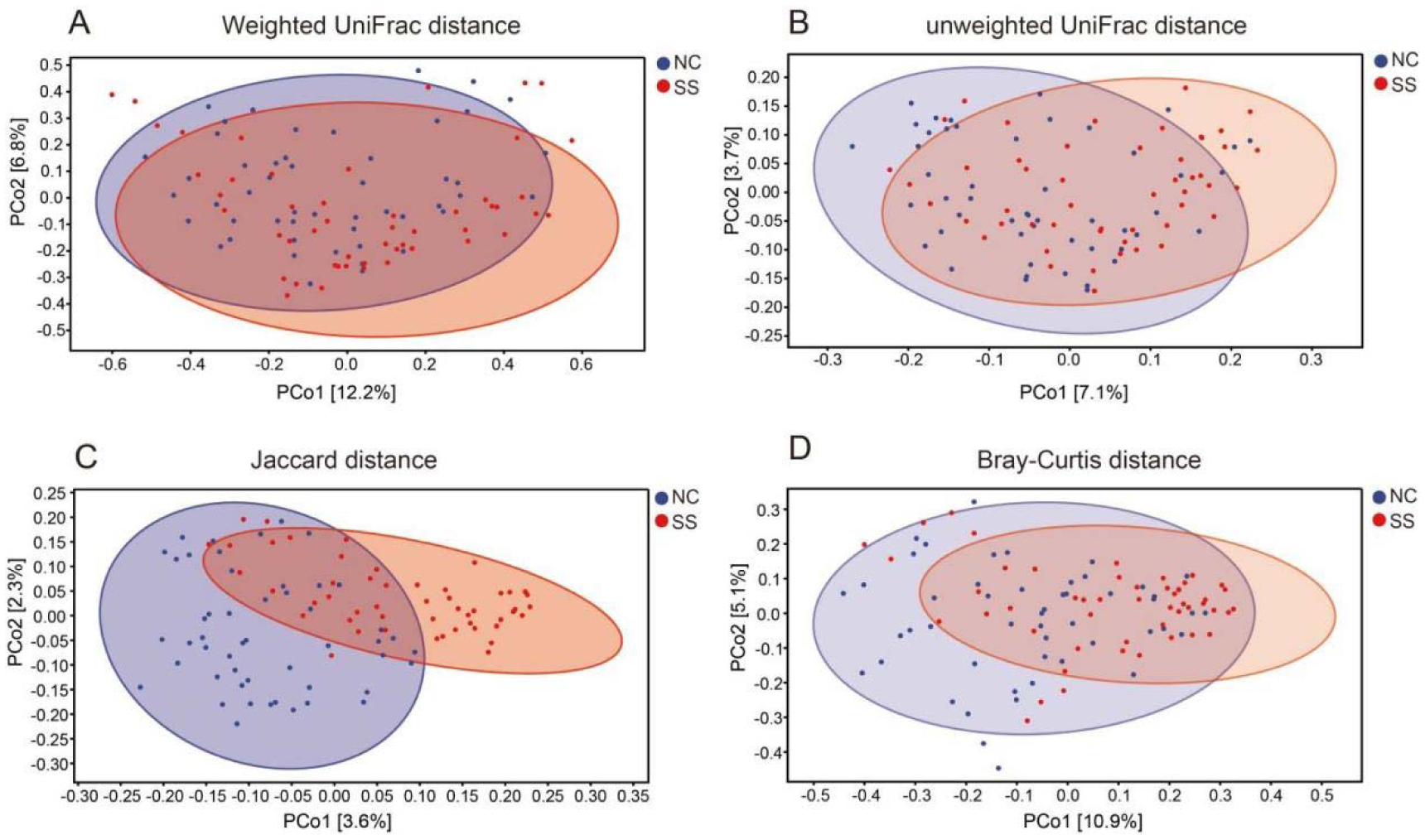
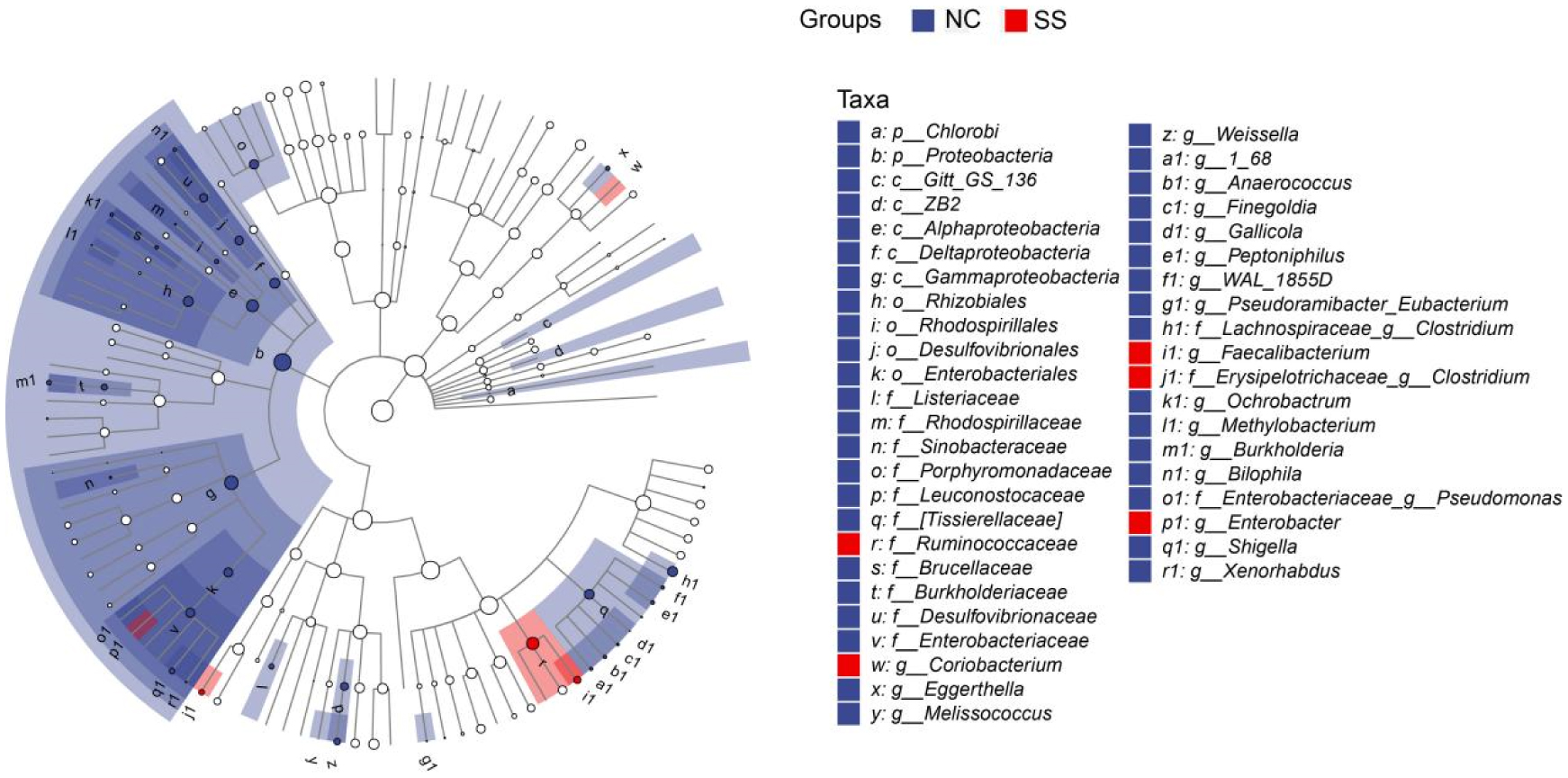
Physical exercise contributes to good health and affects gut microbiota. We investigated the gut microbiota features of students majoring in sports. Fecal samples were collected from the sports majors (SS group, 50 students, 11 males, and 39 females, 18.7 ± 0.8 years old) and non-sports majors (NC group, 50 students, 23 males, and 27 females, 19.0 ± 1.0 years old) for high-throughput sequencing analysis. Their clinical data were statistically analyzed. The results showed that the gut microbiota compositions are similar in evolution and development, and significant differences were observed in the proportion of rare species, such as Fusobacterium, Mitsuokella, Acidaminococcus, and Butyricicoccus. Compared with the NC group, the SS group exhibited a more diverse gut microbiota, with a high abundance of Alistipes, Faecalibacterium, Roseburia, Prevotella, etc., at the genus level, as well as Faecalibacterium prausnitzii, Bifidobacterium adolescentis, Bacteroides uniformis, and Bacteroides plebeius at the species level. The research shows that long-term physical exercise or training can enhance gut microbiota diversity and boost beneficial bacteria, which are conducive to intestinal environmental conditions.
Citation: Benlong Ma, Yifang Zhang, Wenbo Yuan, Nan Hu, Shanshan Li. Gut microbiota differences between sports and non-sports major students: a population-controlled trial[J]. AIMS Bioengineering, 2024, 11(3): 478-488. doi: 10.3934/bioeng.2024021
Physical exercise contributes to good health and affects gut microbiota. We investigated the gut microbiota features of students majoring in sports. Fecal samples were collected from the sports majors (SS group, 50 students, 11 males, and 39 females, 18.7 ± 0.8 years old) and non-sports majors (NC group, 50 students, 23 males, and 27 females, 19.0 ± 1.0 years old) for high-throughput sequencing analysis. Their clinical data were statistically analyzed. The results showed that the gut microbiota compositions are similar in evolution and development, and significant differences were observed in the proportion of rare species, such as Fusobacterium, Mitsuokella, Acidaminococcus, and Butyricicoccus. Compared with the NC group, the SS group exhibited a more diverse gut microbiota, with a high abundance of Alistipes, Faecalibacterium, Roseburia, Prevotella, etc., at the genus level, as well as Faecalibacterium prausnitzii, Bifidobacterium adolescentis, Bacteroides uniformis, and Bacteroides plebeius at the species level. The research shows that long-term physical exercise or training can enhance gut microbiota diversity and boost beneficial bacteria, which are conducive to intestinal environmental conditions.

| [1] |
Ussar S, Fujisaka S, Kahn CR (2016) Interactions between host genetics and gut microbiome in diabetes and metabolic syndrome. Mol Metab 5: 795-803. https://doi.org/10.1016/j.molmet.2016.07.004 
|
| [2] |
Heiman ML, Greenway FL (2016) A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol Metab 5: 317-320. https://doi.org/10.1016/j.molmet.2016.02.005
|
| [3] |
Clarke SF, Murphy EF, O'Sullivan O, et al. (2014) Exercise and associated dietary extremes impact on gut microbial diversity. Gut 63: 1913-1920. https://doi.org/10.1136/gutjnl-2013-306541
|
| [4] |
Campbell SC, Wisniewski PJ, Noji M, et al. (2016) The effect of diet and exercise on intestinal integrity and microbial diversity in mice. PloS One 11: e0150502. https://doi.org/10.1371/journal.pone.0150502
|
| [5] |
Allen JM, Mailing LJ, Niemiro GM, et al. (2018) Exercise alters gut microbiota composition and function in lean and obese humans. Med Sci Sport Exer 50: 747-757. https://doi.org/10.1249/MSS.0000000000001495
|
| [6] |
Campbell SC, Wisniewski PJ (2017) Exercise is a novel promoter of intestinal health and microbial diversity. Exerc Sport Sci Rev 45: 41-47. https://doi.org/10.1249/JES.0000000000000096
|
| [7] |
Kulecka M, Fraczek B, Mikula M, et al. (2020) The composition and richness of the gut microbiota differentiate the top Polish endurance athletes from sedentary controls. Gut Microbes 11: 1374-1384. https://doi.org/10.1080/19490976.2020.1758009
|
| [8] |
Li SS, Qi YL, Chen LX, et al. (2019) Effects of Panax ginseng polysaccharides on the gut microbiota in mice with antibiotic-associated diarrhea. Int J Biol Macromol 124: 931-937. https://doi.org/10.1016/j.ijbiomac.2018.11.271
|
| [9] |
Heck KL, Van Belle G, Simberloff D (1975) Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 56: 1459-1461. https://doi.org/10.2307/1934716
|
| [10] |
Kemp PF, Aller JY (2004) Bacterial diversity in aquatic and other environments: What 16S rDNA libraries can tell us. FEMS Microbiol Ecol 47: 161-177. https://doi.org/10.1016/S0168-6496(03)00257-5
|
| [11] |
Donati Zeppa S, Agostini D, Gervasi M, et al. (2019) Mutual interactions among exercise, sport supplements and microbiota. Nutrients 12: 17. https://doi.org/10.3390/nu12010017
|
| [12] |
Wegierska AE, Charitos IA, Topi S, et al. (2022) The connection between physical exercise and gut microbiota: implications for competitive sports athletes. Sports Med 52: 2355-2369. https://doi.org/10.1007/s40279-022-01696-x
|
| [13] |
Mancin L, Rollo I, Mota JF, et al. (2021) Optimizing microbiota profiles for athletes. Exerc Sport Sci Rev 49: 42-49. https://doi.org/10.1249/JES.0000000000000236
|
| [14] |
Aya JV, Vega LC, Muñoz E, et al. (2024) Divergent gut microbiota: archaeal and bacterial signatures unveil unique patterns in Colombian cyclists compared to weightlifters and non-athletes. Adv Biol 8: 2400069. https://doi.org/10.1002/adbi.202400069
|
| [15] | Lambeth SM, Carson T, Lowe J, et al. (2015) Composition, diversity and abundance of gut microbiome in prediabetes and type 2 diabetes. J Diabetes Obes 2: 1-7. https://doi.org/10.15436/2376-0949.15.031 |
| [16] |
Scher JU, Ubeda C, Artacho A, et al. (2015) Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol 67: 128-139. https://doi.org/10.1002/art.38892
|
| [17] |
Sommer F, Anderson JM, Bharti R, et al. (2017) The resilience of the intestinal microbiota influences health and disease. Nat Rev Microbiol 15: 630-638. https://doi.org/10.1038/nrmicro.2017.58
|
| [18] |
Benevides L, Burman S, Martin R, et al. (2017) New insights into the diversity of the genus Faecalibacterium. Front Microbiol 8: 1790. https://doi.org/10.3389/fmicb.2017.01790
|
| [19] |
Trefflich I, Dietrich S, Braune A, et al. (2021) Short- and branched-chain fatty acids as fecal markers for microbiota activity in vegans and omnivores. Nutrients 13: 1808. https://doi.org/10.3390/nu13061808
|
| [20] |
Hu W, Gao W, Liu Z, et al. (2022) Biodiversity and physiological characteristics of novel faecalibacterium prausnitzii strains isolated from human feces. Microorganisms 10: 297. https://doi.org/10.3390/microorganisms10020297
|
| [21] |
Auger S, Kropp C, Borras-Nogues E, et al. (2022) Intraspecific diversity of microbial anti-inflammatory molecule (MAM) from faecalibacterium prausnitzii. Int J Mol Sci 23: 1705. https://doi.org/10.3390/ijms23031705
|
| [22] |
Ding M, Yang B, Khine WWT, et al. (2021) The species-level composition of the fecal bifidobacterium and lactobacillus genera in Indonesian children differs from that of their mothers. Microorganisms 9: 1995. https://doi.org/10.3390/microorganisms9091995
|
| [23] |
Drago L (2019) Prevotella copri and microbiota in rheumatoid arthritis: fully convincing evidence?. J Clin Med 8: 1837. https://doi.org/10.3390/jcm8111837
|
| [24] |
Khaledi M, Darvishi M, Sameni F, et al. (2024) Association between exercise and changes in gut microbiota profile: a review. Sport Sci Health 20: 273-286. https://doi.org/10.3390/jcm8111837
|
| [25] | Pérez-Castillo IM, Sabag-Daigle A, López-Chicharro J, et al. (2024) The athlete gut microbiota: state of the art and practical guidance. Benef Microbes 26: 97-126. https://doi.org/10.1163/18762891-bja00007 |
| [26] | Mokarrami A, Capacci A, Trio B, et al. (2024) Relationship between gut-microbiota and sport activity. Cent Eur J Sport Sci Med 45: 25-53. https://doi.org/10.18276/cej.2024.1-03 |
| [27] |
Mankowska K, Marchelek-Myśliwiec M, Kochan P, et al. (2022) Microbiota in sports. Arch Microbiol 204: 485. https://doi.org/10.1007/s00203-022-03111-5
|
| [28] |
Zhang YW, Cao MM, Li YJ, et al. (2022) Fecal microbiota transplantation ameliorates bone loss in mice with ovariectomy-induced osteoporosis via modulating gut microbiota and metabolic function. J Orthop Translat 37: 46-60. https://doi.org/10.1016/j.jot.2022.08.003
|
| [29] |
Zhang YW, Cao MM, Li YJ, et al. (2022) A narrative review of the moderating effects and repercussion of exercise intervention on osteoporosis: ingenious involvement of gut microbiota and its metabolites. J Transl Med 20: 490. https://doi.org/10.1186/s12967-022-03700-4
|
| [30] |
Zhang YW, Cao MM, Li YJ, et al. (2023) The regulative effect and repercussion of probiotics and prebiotics on osteoporosis: involvement of brain-gut-bone axis. Crit Rev Food Sci Nutr 63: 7510-7528. https://doi.org/10.1080/10408398.2022.2047005
|
Figures(5) / Tables(1)

Benlong Ma, Yifang Zhang, Wenbo Yuan, Nan Hu, Shanshan Li. Gut microbiota differences between sports and non-sports major students: a population-controlled trial[J]. AIMS Bioengineering, 2024, 11(3): 478-488. doi: 10.3934/bioeng.2024021







 DownLoad:
DownLoad: