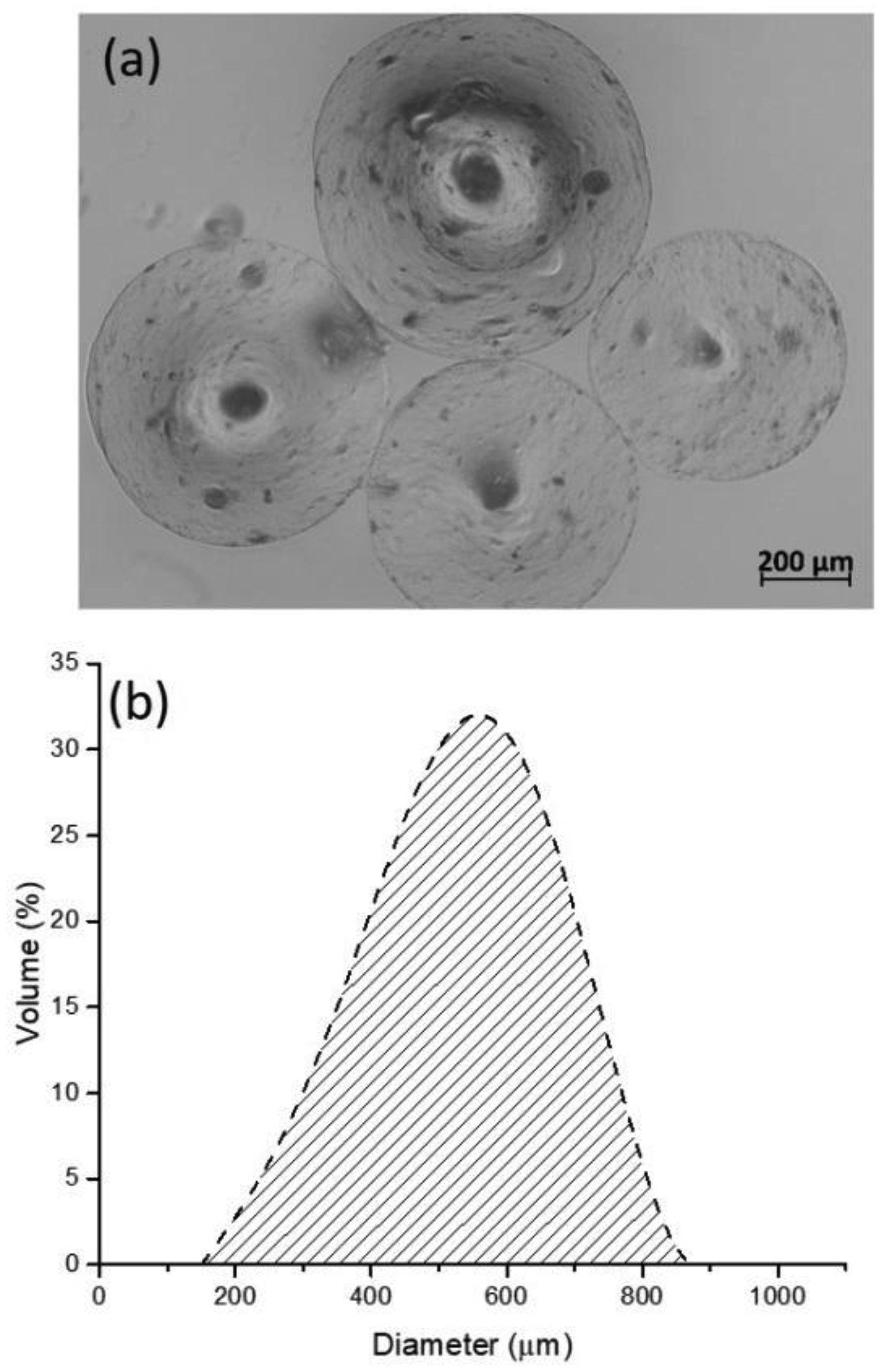
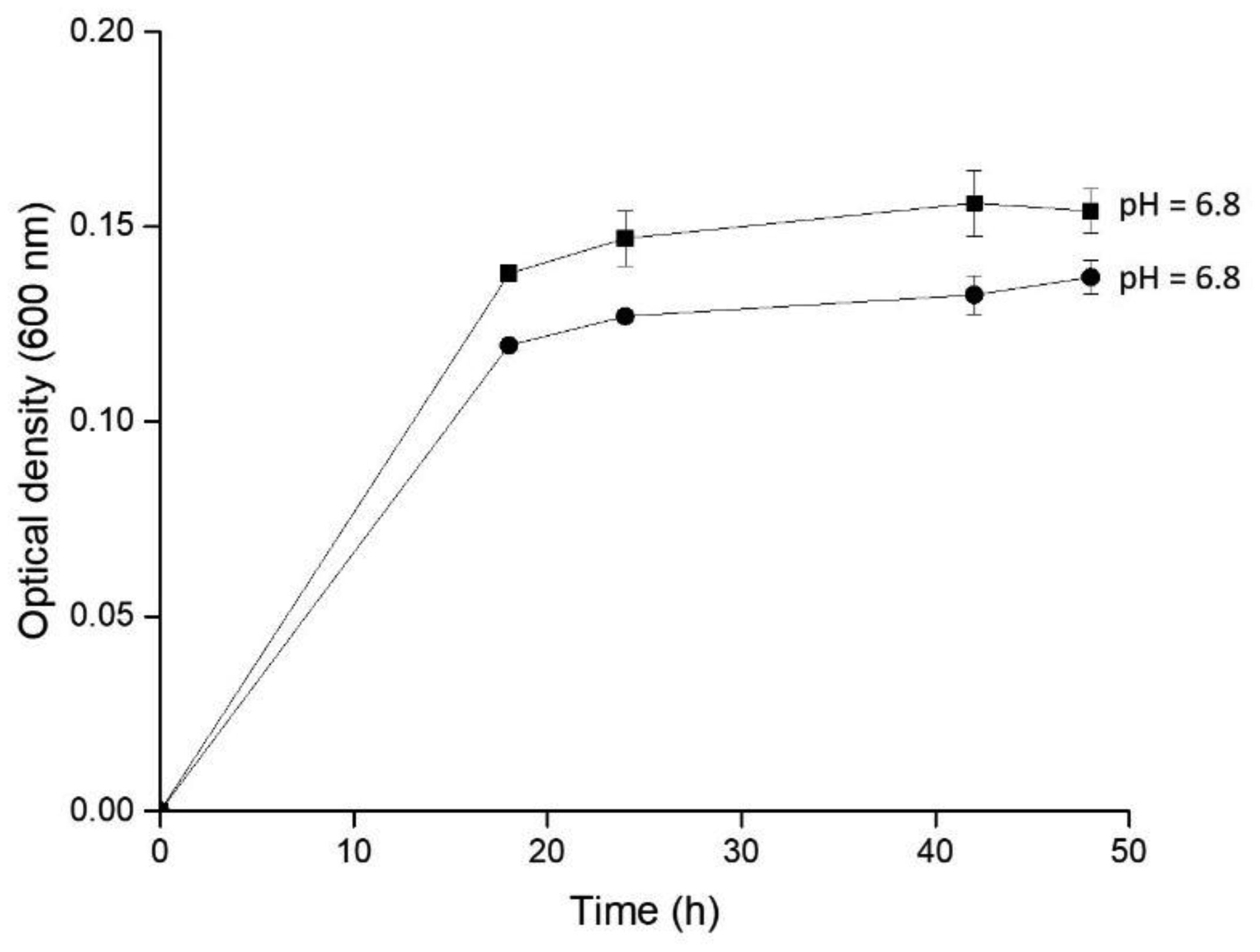
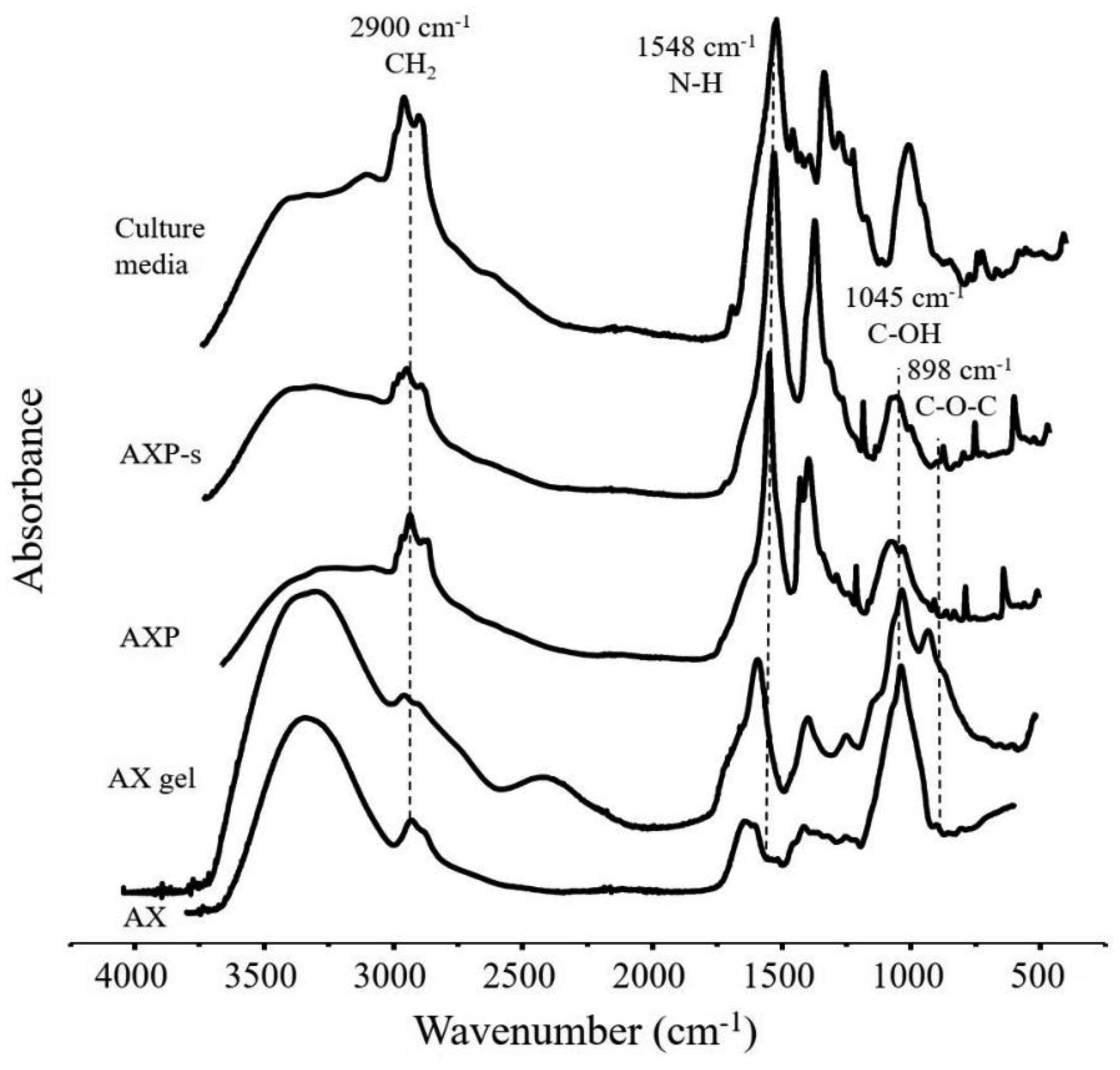
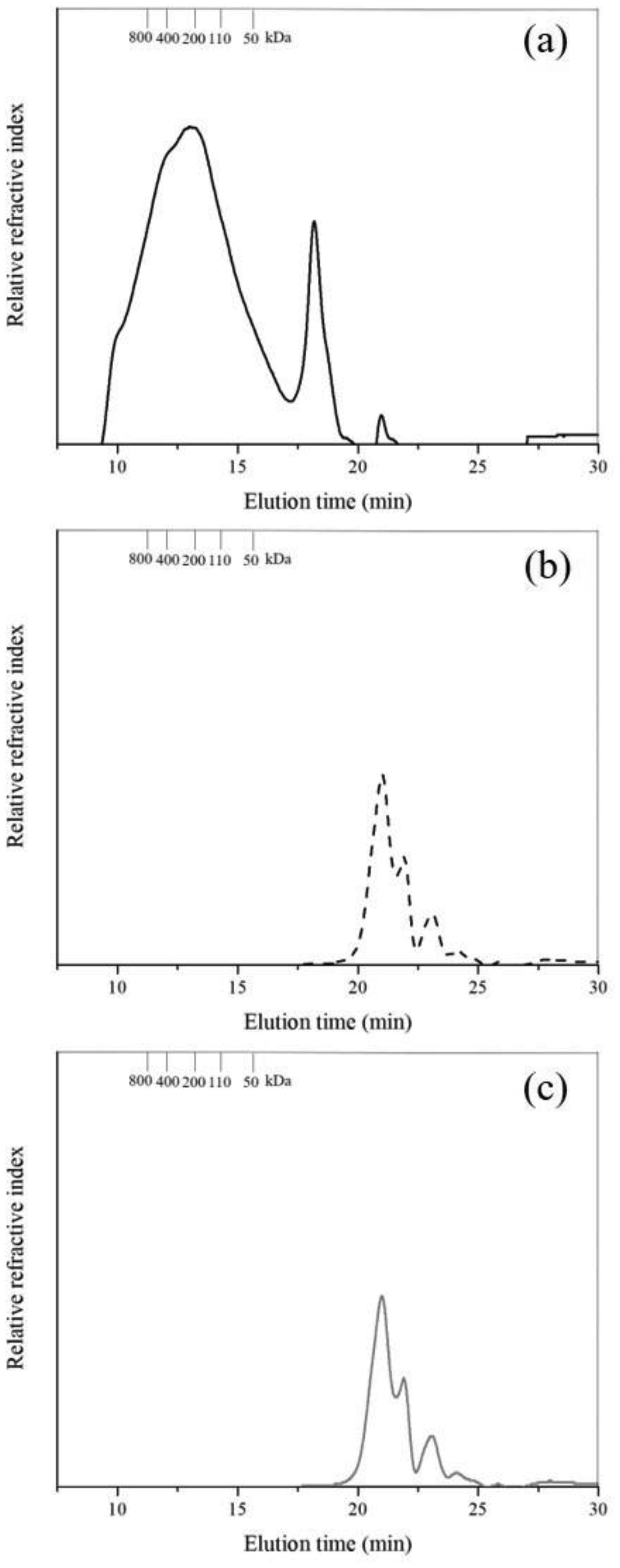
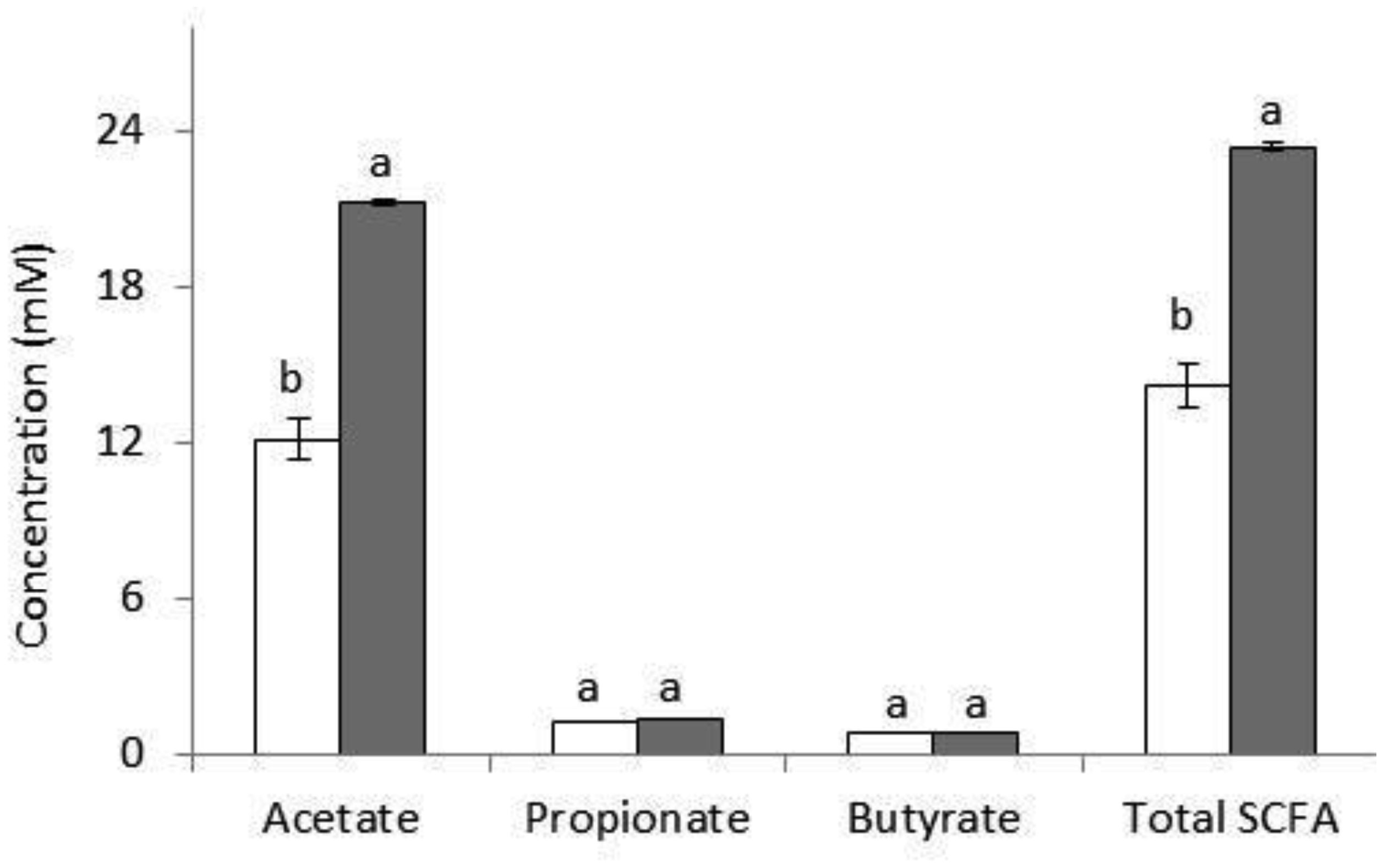
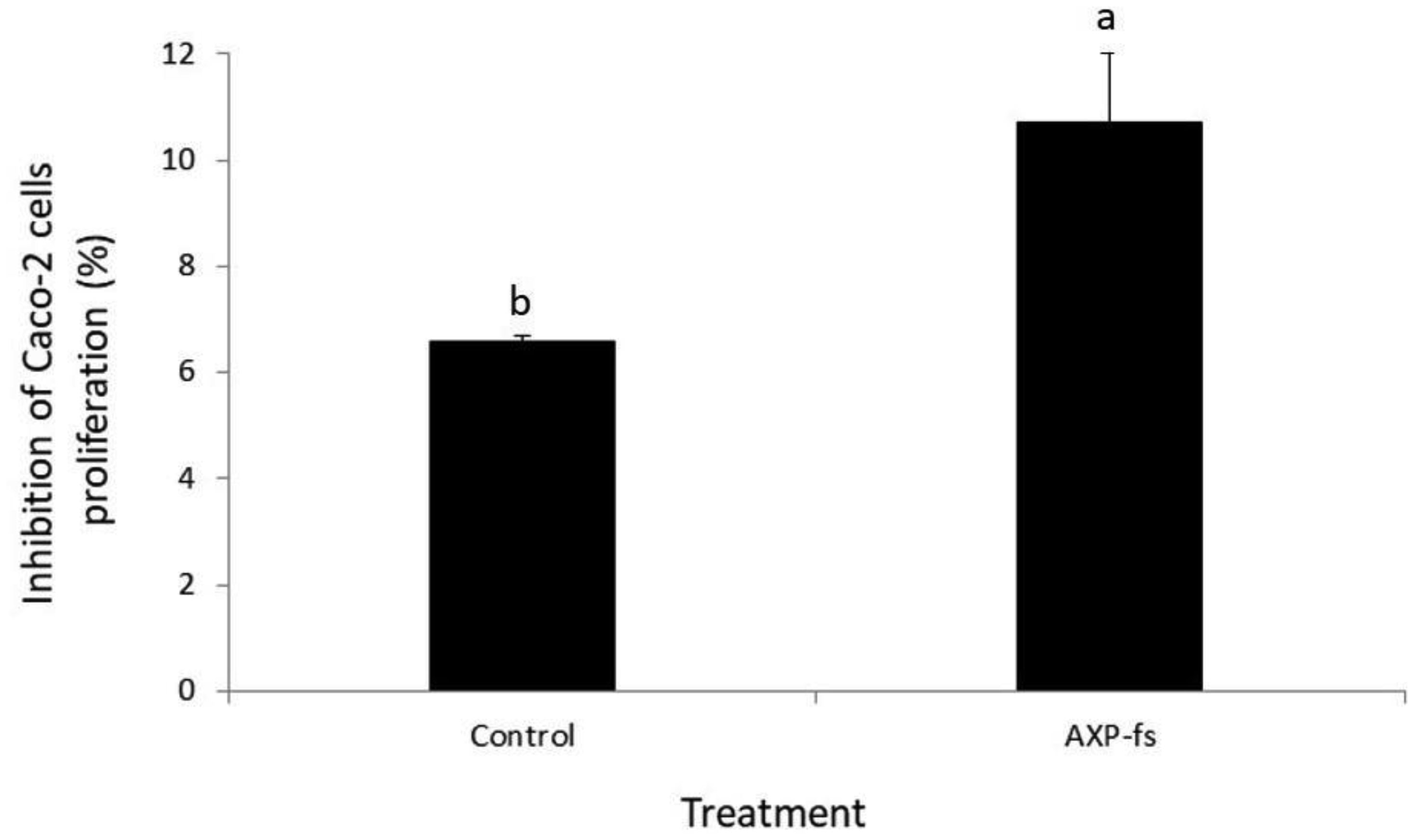
Arabinoxylans (AX) are gelling polysaccharides with potential applications as colon-targeted biomaterials. Nevertheless, the fermentation of highly cross-linked AX particles (AXP) by colonic bacteria and the effect of its fermentation supernatants on the proliferation of human colon cancer cells have not been investigated so far. In this study, electrosprayed AXP were fermented by Bifidobacterium longum, Bifidobacterium adolescentis, and Bacteroides ovatus. The effect of AXP fermentation supernatant (AXP-fs) on the inhibition of the human colon cancer cell line Caco-2 proliferation was investigated. AXP presented a mean diameter of 533 µm, a spherical shape, and a cross-linking content (dimers and trimers of ferulic acid) of 1.65 µg/mg polysaccharide. After 48 h of bacteria exposure, AXP were only partly fermented, probably due to polymeric network steric hindrance that limits the access of bacterial enzymes to the polysaccharide target sites. AXP partial fermentation was evidenced by a moderate short-chain fatty acid production (SCFA) (23 mM) and a collapsed and disintegrated microstructure revealed by scanning electron microscopy. AXP-fs exerted slight inhibition of Caco-2 cell proliferation (11%), which could be attributed to the SCFA generated during partly polysaccharide fermentation. These findings indicate that electrosprayed AXP are a slow-fermentable biomaterial presenting slight anti-cancer properties and potential application in colon cancer prevention.
Citation: Mayra A. Mendez-Encinas, Dora E. Valencia-Rivera, Elizabeth Carvajal-Millan, Humberto Astiazaran-Garcia, Agustín Rascón-Chu, Francisco Brown-Bojorquez. Electrosprayed highly cross-linked arabinoxylan particles: effect of partly fermentation on the inhibition of Caco-2 cells proliferation[J]. AIMS Bioengineering, 2021, 8(1): 52-70. doi: 10.3934/bioeng.2021006
Arabinoxylans (AX) are gelling polysaccharides with potential applications as colon-targeted biomaterials. Nevertheless, the fermentation of highly cross-linked AX particles (AXP) by colonic bacteria and the effect of its fermentation supernatants on the proliferation of human colon cancer cells have not been investigated so far. In this study, electrosprayed AXP were fermented by Bifidobacterium longum, Bifidobacterium adolescentis, and Bacteroides ovatus. The effect of AXP fermentation supernatant (AXP-fs) on the inhibition of the human colon cancer cell line Caco-2 proliferation was investigated. AXP presented a mean diameter of 533 µm, a spherical shape, and a cross-linking content (dimers and trimers of ferulic acid) of 1.65 µg/mg polysaccharide. After 48 h of bacteria exposure, AXP were only partly fermented, probably due to polymeric network steric hindrance that limits the access of bacterial enzymes to the polysaccharide target sites. AXP partial fermentation was evidenced by a moderate short-chain fatty acid production (SCFA) (23 mM) and a collapsed and disintegrated microstructure revealed by scanning electron microscopy. AXP-fs exerted slight inhibition of Caco-2 cell proliferation (11%), which could be attributed to the SCFA generated during partly polysaccharide fermentation. These findings indicate that electrosprayed AXP are a slow-fermentable biomaterial presenting slight anti-cancer properties and potential application in colon cancer prevention.

| [1] |
Mendez-Encinas M A, Carvajal-Millan E, Rascón-Chu A, et al. (2019) Arabinoxylan-based particles: In vitro antioxidant capacity and cytotoxicity on a human colon cell line. Medicina 55: 349. doi: 10.3390/medicina55070349

|
| [2] |
Hopkins MJ, Englyst HN, Macfarlane S, et al. (2003) Degradation of cross-linked and non-cross-linked arabinoxylans by the intestinal microbiota in children. Appl Environ Microbiol 69: 6354-6360. doi: 10.1128/AEM.69.11.6354-6360.2003
|
| [3] |
Martínez-López A L, Carvajal-Millan E, Micard V, et al. (2016) In vitro degradation of covalently cross-linked arabinoxylan hydrogels by bifidobacteria. Carbohydr Polym 144: 76-82. doi: 10.1016/j.carbpol.2016.02.031
|
| [4] |
Paz-Samaniego R, Rascón-Chu A, Brown-Bojorquez F, et al. (2018) Electrospray-assisted fabrication of core-shell arabinoxylan gel particles for insulin and probiotics entrapment. J Appl Polym Sci 135: 46411. doi: 10.1002/app.46411
|
| [5] |
Martínez-López AL, Carvajal-Millan E, Sotelo-Cruz N, et al. (2019) Enzymatically cross-linked arabinoxylan microspheres as oral insulin delivery system. Int J Biol Macromol 126: 952-959. doi: 10.1016/j.ijbiomac.2018.12.192
|
| [6] |
Carvajal-Millan E, Vargas-Albores F, Fierro-Islas JM, et al. (2020) Arabinoxylans and gelled arabinoxylans used as anti-obesogenic agents could protect the stability of intestinal microbiota of rats consuming high-fat diets. Int J Food Sci Nutr 71: 74-83. doi: 10.1080/09637486.2019.1610729
|
| [7] |
Izydorczyk MS, Biliaderis CG (1995) Cereal arabinoxylans: advances in structure and physicochemical properties. Carbohydr Polym 28: 33-48. doi: 10.1016/0144-8617(95)00077-1
|
| [8] |
Smith MM, Hartley RD (1983) Occurrence and nature of ferulic acid substitution of cell-wall polysaccharides in graminaceous plants. Carbohydr Res 118: 65-80. doi: 10.1016/0008-6215(83)88036-7
|
| [9] |
Martínez-López AL, Carvajal-Millan E, Marquez-Escalante J, et al. (2019) Enzymatic cross-linking of ferulated arabinoxylan: effect of laccase or peroxidase catalysis on the gel characteristics. Food Sci Biotechnol 28: 311-318. doi: 10.1007/s10068-018-0488-9
|
| [10] |
Figueroa-Espinoza MC, Morel MH, Surget A, et al. (1999) Oxidative cross-linking of wheat arabinoxylans by manganese peroxidase. Comparison with laccase and horseradish peroxidase. Effect of cysteine and tyrosine on gelation. J Sci Food Agric 79: 460-463. doi: 10.1002/(SICI)1097-0010(19990301)79:3<460::AID-JSFA268>3.0.CO;2-7
|
| [11] |
Carvajal-Millan E, Guigliarelli B, Belle V, et al. (2005) Storage stability of laccase induced arabinoxylan gels. Carbohydr Polym 59: 181-188. doi: 10.1016/j.carbpol.2004.09.008
|
| [12] |
Morales-Burgos A M, Carvajal-Millan E, Rascón-Chu A, et al. (2019) Tailoring reversible insulin aggregates loaded in electrosprayed arabinoxylan microspheres intended for colon-targeted delivery. J Appl Polym Sci 136: 47960. doi: 10.1002/app.47960
|
| [13] |
Berlanga-Reyes CM, Carvajal-Millán E, Lizardi-Mendoza J, et al. (2009) Maize arabinoxylan gels as protein delivery matrices. Molecules 14: 1475-1482. doi: 10.3390/molecules14041475
|
| [14] |
Hernández-Espinoza AB, Piñón-Muñiz MI, Rascón-Chu A, et al. (2012) Lycopene/arabinoxylan gels: Rheological and controlled release characteristics. Molecules 17: 2428-2436. doi: 10.3390/molecules17032428
|
| [15] |
Paz-Samaniego R, Carvajal-Millan E, Sotelo-Cruz N, et al. (2016) Maize processing waste water upcycling in Mexico: Recovery of arabinoxylans for probiotic encapsulation. Sustainability 8: 1104. doi: 10.3390/su8111104
|
| [16] |
Chen Z, Li S, Fu Y, et al. (2019) Arabinoxylan structural characteristics, interaction with gut microbiota and potential health functions. J Funct Foods 54: 536-551. doi: 10.1016/j.jff.2019.02.007
|
| [17] |
Morrison DJ, Preston T (2016) Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7: 189-200. doi: 10.1080/19490976.2015.1134082
|
| [18] |
Ohara T, Mori T (2019) Antiproliferative effects of short-chain fatty acids on human colorectal cancer cells via gene expression Inhibition. Anticancer Res 39: 4659-4666. doi: 10.21873/anticanres.13647
|
| [19] |
Marques C, Oliveira CSF, Alves S, et al. (2013) Acetate-induced apoptosis in colorectal carcinoma cells involves lysosomal membrane permeabilization and cathepsin D release. Cell Death Dis 4: e507. doi: 10.1038/cddis.2013.29
|
| [20] |
Glei M, Hofmann T, Küster K, et al. (2006) Both wheat (Triticum aestivum) bran arabinoxylans and gut flora-mediated fermentation products protect human colon cells from genotoxic activities of 4-hydroxynonenal and hydrogen peroxide. J Agric Food Chem 54: 2088-2095. doi: 10.1021/jf052768e
|
| [21] | Roy N, Narayanankutty A, Nazeem PA, et al. (2016) Plant phenolics ferulic acid and p-coumaric acid inhibit colorectal cancer cell proliferation through EGFR down- regulation. Asian Pacific J Cancer Prev 17: 4019-4023. |
| [22] |
Janicke B, Önning G, Oredsson SM (2005) Differential effects of ferulic acid and p-coumaric acid on S phase distribution and length of S phase in the human colonic cell line Caco-2. J Agric Food Chem 53: 6658-6665. doi: 10.1021/jf050489l
|
| [23] |
Janicke B, Hegardt C, Krogh M, et al. (2011) The antiproliferative effect of dietary fiber phenolic compounds ferulic acid and p-coumaric acid on the cell cycle of Caco-2 cells. Nutr Cancer 63: 611-622. doi: 10.1080/01635581.2011.538486
|
| [24] | Rosa LS, Silva NJA, Soares NCP, et al. (2016) Anticancer properties of phenolic acids in colon cancer–a review. J Nutr Food Sci 6: 468. |
| [25] |
Hughes SA, Shewry PR, Li L, et al. (2007) In vitro fermentation by human fecal microflora of wheat arabinoxylans. J Agric Food Chem 55: 4589-4595. doi: 10.1021/jf070293g
|
| [26] |
Pastell H, Westermann P, Meyer AS, et al. (2009) In vitro fermentation of arabinoxylan-derived carbohydrates by bifidobacteria and mixed fecal microbiota. J Agric Food Chem 57: 8598-8606. doi: 10.1021/jf901397b
|
| [27] |
Snelders J, Olaerts H, Dornez E, et al. (2014) Structural features and feruloylation modulate the fermentability and evolution of antioxidant properties of arabinoxylanoligosaccharides during in vitro fermentation by human gut derived microbiota. J Funct Foods 10: 1-12. doi: 10.1016/j.jff.2014.05.011
|
| [28] |
De Anda-Flores Y, Carvajal-Millan E, Lizardi-Mendoza J, et al. (2020) Covalently cross-linked nanoparticles based on ferulated arabinoxylans recovered from a distiller's dried grains byproduct. Processes 8: 691. doi: 10.3390/pr8060691
|
| [29] |
Rascón-Chu A, Díaz-Baca JA, Carvajal-Millan E, et al. (2018) Electrosprayed core–shell composite microbeads based on pectin-arabinoxylans for insulin carrying: aggregation and size dispersion control. Polymers 10: 108. doi: 10.3390/polym10020108
|
| [30] |
Rascón-Chu A, Martínez-López AL, Berlanga-Reyes C, et al. (2012) Arabinoxylans gels as lycopene carriers: In vitro degradation by colonic bacteria. Mater Res Soc Symp Proc 1487: 26-32. doi: 10.1557/opl.2012.1527
|
| [31] |
Mendez-Encinas MA, Carvajal-Millan E, Yadav MP, et al. (2019) Partial removal of protein associated with arabinoxylans: Impact on the viscoelasticity, crosslinking content, and microstructure of the gels formed. J Appl Polym Sci 136: 47300. doi: 10.1002/app.47300
|
| [32] |
Van Laere KMJ, Hartemink R, Bosveld M, et al. (2000) Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J Agric Food Chem 48: 1644-1652. doi: 10.1021/jf990519i
|
| [33] |
Crittenden R, Karppinen S, Ojanen S, et al. (2002) In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria. J Sci Food Agric 82: 781-789. doi: 10.1002/jsfa.1095
|
| [34] |
Zhao G, Nyman M, Åke Jönsson J (2006) Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed Chromatogr 20: 674-682. doi: 10.1002/bmc.580
|
| [35] |
Chen L, Lin X, Teng H (2020) Emulsions loaded with dihydromyricetin enhance its transport through Caco-2 monolayer and improve anti-diabetic effect in insulin resistant HepG2 cell. J Funct Foods 64: 103672. doi: 10.1016/j.jff.2019.103672
|
| [36] |
Hernandez J, Goycoolea FM, Quintero J, et al. (2007) Sonoran propolis: Chemical composition and antiproliferative activity on cancer cell lines. Planta Med 73: 1469-1474. doi: 10.1055/s-2007-990244
|
| [37] |
Martínez-López AL, Carvajal-Millan E, Miki-Yoshida M, et al. (2013) Arabinoxylan microspheres: Structural and textural characteristics. Molecules 18: 4640-4650. doi: 10.3390/molecules18044640
|
| [38] |
Barry JL, Hoebler C, Macfarlane GT, et al. (1995) Estimation of the fermentability of dietary fibre in vitro: a European interlaboratory study. Br J Nutr 74: 303-322. doi: 10.1079/BJN19950137
|
| [39] |
Paesani C, Salvucci E, Moiraghi M, et al. (2019) Arabinoxylan from Argentinian whole wheat flour promote the growth of Lactobacillus reuteri and Bifidobacterium breve. Lett Appl Microbiol 68: 142-148. doi: 10.1111/lam.13097
|
| [40] |
Rivière A, Moens F, Selak M, et al. (2014) The ability of bifidobacteria to degrade arabinoxylan oligosaccharide constituents and derived oligosaccharides is strain dependent. Appl Environ Microbiol 80: 204-217. doi: 10.1128/AEM.02853-13
|
| [41] |
Pollet A, Van Craeyveld V, Van de Wiele T, et al. (2012) In vitro fermentation of arabinoxylan oligosaccharides and low molecular mass arabinoxylans with different structural properties from wheat (Triticum aestivum L.) bran and psyllium (Plantago ovata Forsk) seed husk. J Agric Food Chem 60: 946-954. doi: 10.1021/jf203820j
|
| [42] |
Feng G, Flanagan BM, Mikkelsen D, et al. (2018) Mechanisms of utilisation of arabinoxylans by a porcine faecal inoculum: competition and co-operation. Sci Rep 8: 1-11. doi: 10.1038/s41598-017-17765-5
|
| [43] |
Martínez-López AL, Carvajal-Millan E, López-Franco YL, et al. (2014) Antioxidant activity of maize bran arabinoxylan microspheres. Food Composition and Analysis: Methods and Strategies Toronto: CRC Press, 19-28. doi: 10.1201/b16843-3
|
| [44] |
Hromádková Z, Paulsen BS, Polovka M, et al. (2013) Structural features of two heteroxylan polysaccharide fractions from wheat bran with anti-complementary and antioxidant activities. Carbohydr Polym 93: 22-30. doi: 10.1016/j.carbpol.2012.05.021
|
| [45] |
Morales-Ortega A, Carvajal-Millan E, López-Franco Y, et al. (2013) Characterization of water extractable arabinoxylans from a spring wheat flour: rheological properties and microstructure. Molecules 18: 8417-8428. doi: 10.3390/molecules18078417
|
| [46] |
Stepan AM, Eceiza A, Toriz G, et al. (2014) Corncob arabinoxylan for new materials. Carbohydr Polym 102: 12-20. doi: 10.1016/j.carbpol.2013.11.011
|
| [47] |
Urias-Orona V, Huerta-Oros J, Carvajal-Millán E, et al. (2010) Component analysis and free radicals scavenging activity of Cicer arietinum L. husk pectin. Molecules 15: 6948-6955. doi: 10.3390/molecules15106948
|
| [48] |
Ou J, Sun Z (2014) Feruloylated oligosaccharides: structure, metabolism and function. J Funct Foods 7: 90-100. doi: 10.1016/j.jff.2013.09.028
|
| [49] |
Snelders J, Dornez E, Delcour JA, et al. (2013) Ferulic acid content and appearance determine the antioxidant capacity of arabinoxylanoligosaccharides. J Agric Food Chem 61: 10173-10182. doi: 10.1021/jf403160x
|
| [50] |
Hespell RB, O'Bryan PJ (1992) Purification and characterization of an α-L-arabinofuranosidase from Butyrivibrio fibrisolvens GS113. Appl Environ Microbiol 58: 1082-1088. doi: 10.1128/AEM.58.4.1082-1088.1992
|
| [51] |
Macy JM, Ljungdahl LG, Gottschalk G (1978) Pathway of succinate and propionate formation in Bacteroides fragilis. J Bacteriol 134: 84-91. doi: 10.1128/JB.134.1.84-91.1978
|
| [52] |
Sadeghi Ekbatan S, Li XQ, Ghorbani M, et al. (2018) Chlorogenic acid and its microbial metabolites exert anti-proliferative effects, S-phase cell-cycle arrest and apoptosis in human colon cancer Caco-2 cells. Int J Mol Sci 19: 723. doi: 10.3390/ijms19030723
|
| [53] |
Louis P, Hold GL, Flint HJ (2014) The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol 12: 661-672. doi: 10.1038/nrmicro3344
|
| [54] | Mendez-Encinas MA, Carvajal-Millan E, Rascon-Chu A, et al. (2018) Ferulated arabinoxylans and their gels: Functional properties and potential application as antioxidant and anticancer agent. Oxid Med Cell Longev 2018: 2314759. |
| [55] |
Ríos-Covián D, Ruas-Madiedo P, Margolles A, et al. (2016) Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol 7: 185. doi: 10.3389/fmicb.2016.00185
|
| [56] |
Rivière A, Selak M, Lantin D, et al. (2016) Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front Microbiol 7: 979. doi: 10.3389/fmicb.2016.00979
|
| [57] |
Longley DB, Harkin DP, Johnston PG (2003) 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat Rev Cancer 3: 330-338. doi: 10.1038/nrc1074
|

Figures(7) / Tables(1)

Mayra A. Mendez-Encinas, Dora E. Valencia-Rivera, Elizabeth Carvajal-Millan, Humberto Astiazaran-Garcia, Agustín Rascón-Chu, Francisco Brown-Bojorquez. Electrosprayed highly cross-linked arabinoxylan particles: effect of partly fermentation on the inhibition of Caco-2 cells proliferation[J]. AIMS Bioengineering, 2021, 8(1): 52-70. doi: 10.3934/bioeng.2021006







 DownLoad:
DownLoad: