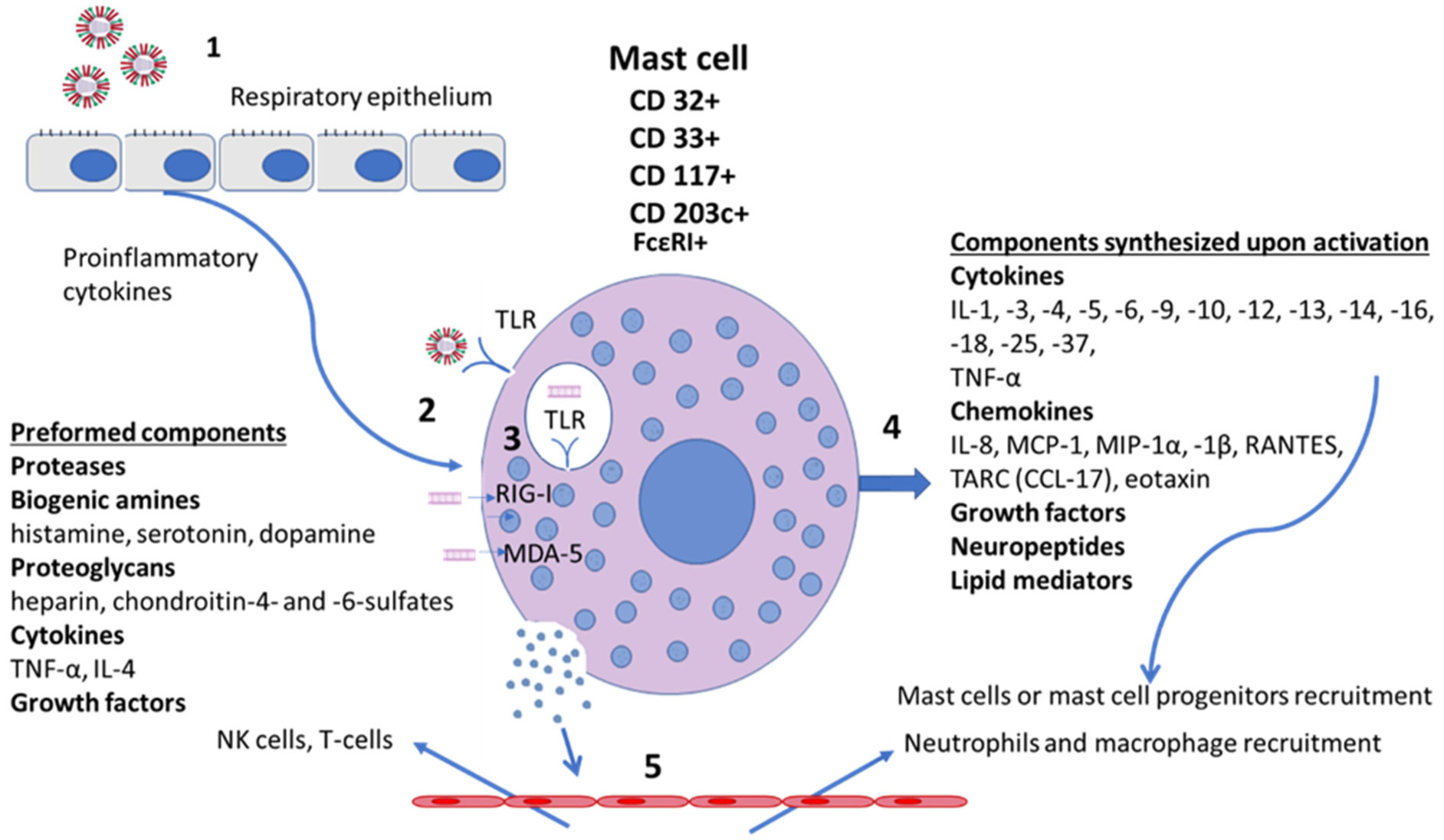
Mast cells (MCs) are a part of the innate immune system and express receptors for microbial and viral pathogens characteristic of this system. The pathological role of MCs has been demonstrated for a number of highly virulent viral infections. The role of MCs and their Fc receptors for IgE in the immediate-type hypersensitivity reactions and in immunocomplex reactions is well-known, although the role of MCs and their Fc receptors for IgG (FcγR) in immunocomplex processes is much less studied. Antibody-dependent enhancement syndrome (ADE) has been observed in a number of viral infections and is associated with greater secondary infection. ADE is enhanced by virus-specific antibodies, which are not involved in the virus penetration into the cell but are capable of forming immune complexes. The role of MCs in ADE is well-established for dengue infection, RSV infection and coronavirus (CoV) infection. The involvement of IgG-mediated mast cell responses in other human viral infections including Coronavirus disease 2019 (COVID-19) is poorly understood. Recently discovered mast cell activation disease is considered one of the causes of severe post-infectious complications in COVID-19. If the role of MCs in the pathogenesis of severe viral infections, including ADE in recurrent viral infection is clarified, these cells and the products they release may serve as promising targets for such therapeutic agents as histamine receptor blockers or membrane stabilizers to prevent possible complications.
Citation: Andrey Mamontov, Alexander Polevshchikov, Yulia Desheva. Mast cells in severe respiratory virus infections: insights for treatment and vaccine administration[J]. AIMS Allergy and Immunology, 2023, 7(1): 1-23. doi: 10.3934/Allergy.2023001
Mast cells (MCs) are a part of the innate immune system and express receptors for microbial and viral pathogens characteristic of this system. The pathological role of MCs has been demonstrated for a number of highly virulent viral infections. The role of MCs and their Fc receptors for IgE in the immediate-type hypersensitivity reactions and in immunocomplex reactions is well-known, although the role of MCs and their Fc receptors for IgG (FcγR) in immunocomplex processes is much less studied. Antibody-dependent enhancement syndrome (ADE) has been observed in a number of viral infections and is associated with greater secondary infection. ADE is enhanced by virus-specific antibodies, which are not involved in the virus penetration into the cell but are capable of forming immune complexes. The role of MCs in ADE is well-established for dengue infection, RSV infection and coronavirus (CoV) infection. The involvement of IgG-mediated mast cell responses in other human viral infections including Coronavirus disease 2019 (COVID-19) is poorly understood. Recently discovered mast cell activation disease is considered one of the causes of severe post-infectious complications in COVID-19. If the role of MCs in the pathogenesis of severe viral infections, including ADE in recurrent viral infection is clarified, these cells and the products they release may serve as promising targets for such therapeutic agents as histamine receptor blockers or membrane stabilizers to prevent possible complications.
antibody-dependent enhancement
antigen presenting cells
central nervous system
coronavirus
coronavirus disease
dengue virus
extracellular vesicles
high-affinity IgE receptor
Fc receptors for IgG
inactivated influenza vaccines
interleukin
mast cell
melanoma differentiation-associated protein 5
mast cells extracellular trap
MCs, containing only tryptase
MCs containing tryptase and chymase
MCs containing only chymase
nucleotide-binding oligomerization domain
Retinoic acid-inducible gene I
respiratory syncytial virus
rhinovirus
severe acute respiratory syndrome
Toll-like receptors
tumor necrosis factor alpha
vaccine-associated enhanced respiratory disease

| [1] | Mahalingam S, Schwarze J, Zaid A, et al. (2006) Perspective on the host response to human metapneumovirus infection: what can we learn from respiratory syncytial virus infections?. Microbes Infect 8: 285-293. https://doi.org/10.1016/j.micinf.2005.07.001 |
| [2] | Liu H, Tan J, Liu J, et al. (2020) Altered mast cell activity in response to rhinovirus infection provides novel insight into asthma. J Asthma 57: 459-467. https://doi.org/10.1080/02770903.2019.1585870 |
| [3] | Vargas SO, Kozakewich HP, Perez-Atayde AR, et al. (2004) Pathology of human metapneumovirus infection: insights into the pathogenesis of a newly identified respiratory virus. Pediatr Devel Pathol 7: 478-486. https://doi.org/10.1007/s10024-004-1011-2 |
| [4] | Pawełczyk M, Kowalski ML (2017) The role of human parainfluenza virus infections in the immunopathology of the respiratory tract. Curr Allergy Asthma Rep 17: 1-10. https://doi.org/10.1007/s11882-017-0685-2 |
| [5] | Krystel-Whittemore M, Dileepan KN, Wood JG (2016) Mast cell: a multi-functional master cell. Front Immunol 6: 620. https://doi.org/10.3389/fimmu.2015.00620 |
| [6] | Xanthos DN, Gaderer S, Drdla R, et al. (2011) Central nervous system mast cells in peripheral inflammatory nociception. Mol Pain 7: 1744-8069. https://doi.org/10.1186/1744-8069-7-42 |
| [7] | Agier J, Różalska S, Wiktorska M, et al. (2018) The RLR/NLR expression and pro-inflammatory activity of tissue mast cells are regulated by cathelicidin LL-37 and defensin hBD-2. Sci Rep 8: 1-16. https://doi.org/10.1038/s41598-018-30289-w |
| [8] | Forsythe P (2019) Mast cells in neuroimmune interactions. Trends Neurosci 42: 43-55. https://doi.org/10.1016/j.tins.2018.09.006 |
| [9] | Erjefält JS (2014) Mast cells in human airways: the culprit?. Eur Respir Rev 23: 299-307. https://doi.org/10.1183/09059180.00005014 |
| [10] | Huber M, Cato ACB, Ainooson GK, et al. (2019) Regulation of the pleiotropic effects of tissue-resident mast cells. J Allergy Clin Immun 144: S31-S45. https://doi.org/10.1016/j.jaci.2019.02.004 |
| [11] | Dawicki W, Marshall JS (2007) New and emerging roles for mast cells in host defence. Curr Opin Immunol 19: 31-38. https://doi.org/10.1016/j.coi.2006.11.006 |
| [12] | Tiligada E, Ennis M (2020) Histamine pharmacology: from Sir Henry Dale to the 21st century. Brit J Pharmacol 177: 469-489. https://doi.org/10.1111/bph.14524 |
| [13] | Hoth M, Penner R (1992) Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature 355: 353-356. https://doi.org/10.1038/355353a0 |
| [14] | Thangam EB, Jemima EA, Singh H, et al. (2018) The role of histamine and histamine receptors in mast cell-mediated allergy and inflammation: the hunt for new therapeutic targets. Front Immunol 9: 1873. https://doi.org/10.3389/fimmu.2018.01873 |
| [15] | Weigand LA, Myers AC, Meeker S, et al. (2009) Mast cell-cholinergic nerve interaction in mouse airways. J Phy 587: 3355-3362. https://doi.org/10.1113/jphysiol.2009.173054 |
| [16] | Wilhelm M, Silver R, Silverman AJ (2005) Central nervous system neurons acquire mast cell products via transgranulation. Eur J Neurosci 22: 2238-2248. https://doi.org/10.1111/j.1460-9568.2005.04429.x |
| [17] | Schiller M, Ben-Shaanan TL, Rolls A (2021) Neuronal regulation of immunity: why, how and where?. Nat Rev Immunol 21: 20-36. https://doi.org/10.1038/s41577-020-0387-1 |
| [18] | Fujita Y, Yoshioka Y, Ito S, et al. (2014) Intercellular communication by extracellular vesicles and their microRNAs in asthma. Clin Ther 36: 873-881. https://doi.org/10.1016/j.clinthera.2014.05.006 |
| [19] | Valadi H, Ekström K, Bossios A, et al. (2007) Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 9: 654-659. https://doi.org/10.1038/ncb1596 |
| [20] | Yin Y, Shelke GV, Lässer C, et al. (2020) Extracellular vesicles from mast cells induce mesenchymal transition in airway epithelial cells. Resp Res 21: 1-13. https://doi.org/10.1186/s12931-020-01346-8 |
| [21] | Marshall JS, Portales-Cervantes L, Leong E (2019) Mast cell responses to viruses and pathogen products. Int J Mol Sci 20: 4241. https://doi.org/10.3390/ijms20174241 |
| [22] | Ding J, Fang Y, Xiang Z (2015) Antigen/IgG immune complex-primed mucosal mast cells mediate antigen-specific activation of co-cultured T cells. Immunology 144: 387-394. https://doi.org/10.1111/imm.12379 |
| [23] | Galli SJ, Tsai M (2012) IgE and mast cells in allergic disease. Nat Med 18: 693-704. https://doi.org/10.1038/nm.2755 |
| [24] | Seneviratne SL, Maitland A, Afrin L (2017) Mast cell disorders in Ehlers–Danlos syndrome. Am J Med Genet 175: 226-236. https://doi.org/10.1002/ajmg.c.31555 |
| [25] | Dudeck A, Köberle M, Goldmann O, et al. (2019) Mast cells as protectors of health. J Allergy Clin Immunol 144: S4-S18. https://doi.org/10.1016/j.jaci.2018.10.054 |
| [26] | Möllerherm H, von Köckritz-Blickwede M, Branitzki-Heinemann K (2016) Antimicrobial activity of mast cells: role and relevance of extracellular DNA traps. Front Immunol 7: 265. https://doi.org/10.3389/fimmu.2016.00265 |
| [27] | Mukai K, Tsai M, Starkl P, et al. (2016) IgE and mast cells in host defense against parasites and venoms. Springer Semin Immun 38: 581-603. https://doi.org/10.1007/s00281-016-0565-1 |
| [28] | Malaviya R, Ross EA, MacGregor JI, et al. (1994) Mast cell phagocytosis of FimH-expressing enterobacteria. J Immunol 152: 1907-1914. |
| [29] | Bruns S, Kniemeyer O, Hasenberg M, et al. (2010) Production of extracellular traps against Aspergillus fumigatus in vitro and in infected lung tissue is dependent on invading neutrophils and influenced by hydrophobin RodA. PLoS Pathog 6: e1000873. https://doi.org/10.1371/journal.ppat.1000873 |
| [30] | Brinkmann V, Zychlinsky A (2007) Beneficial suicide: why neutrophils die to make NETs. Nat Rev Microbiol 5: 577-582. https://doi.org/10.1038/nrmicro1710 |
| [31] | von Köckritz-Blickwede M, Goldmann O, Thulin P, et al. (2008) Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 111: 3070-3080. https://doi.org/10.1182/blood-2007-07-104018 |
| [32] | Lotfi-Emran S, Ward BR, Le QT, et al. (2018) Human mast cells present antigen to autologous CD4+ T cells. J Allergy Clin Immun 141: 311-321. https://doi.org/10.1016/j.jaci.2017.02.048 |
| [33] | Stelekati E, Bahri R, D'Orlando O, et al. (2009) Mast cell-mediated antigen presentation regulates CD8+ T cell effector functions. Immunity 31: 665-676. https://doi.org/10.1016/j.immuni.2009.08.022 |
| [34] | Kambayashi T, Allenspach EJ, Chang JT, et al. (2009) Inducible MHC class II expression by mast cells supports effector and regulatory T cell activation. J Immunol 182: 4686-4695. https://doi.org/10.4049/jimmunol.0803180 |
| [35] | Varricchi G, de Paulis A, Marone G, et al. (2019) Future needs in mast cell biology. Int J Mol Sci 20: 4397. https://doi.org/10.3390/ijms20184397 |
| [36] | Bruhns P, Frémont S, Daëron M (2005) Regulation of allergy by Fc receptors. Curr Opin Immunol 17: 662-669. https://doi.org/10.1016/j.coi.2005.09.012 |
| [37] | Overed-Sayer C, Rapley L, Mustelin T, et al. (2014) Are mast cells instrumental for fibrotic diseases?. Front Pharmacol 4: 174. https://doi.org/10.3389/fphar.2013.00174 |
| [38] | Andersson CK, Andersson-Sjöland A, Mori M, et al. (2011) Activated MCTC mast cells infiltrate diseased lung areas in cystic fibrosis and idiopathic pulmonary fibrosis. Resp Res 12: 1-13. https://doi.org/10.1186/1465-9921-12-139 |
| [39] | Londono-Renteria B, Marinez-Angarita JC, Troupin A, et al. (2017) Role of mast cells in dengue virus pathogenesis. DNA Cell Biol 36: 423-427. https://doi.org/10.1089/dna.2017.3765 |
| [40] | Rathore APS, St John AL (2020) Protective and pathogenic roles for mast cells during viral infections. Curr Opin Immunol 66: 74-81. https://doi.org/10.1016/j.coi.2020.05.003 |
| [41] | Furuta T, Murao LA, Lan NT, et al. (2012) Association of mast cell-derived VEGF and proteases in Dengue shock syndrome. PLoS Negl Trop Dis 6: e1505. https://doi.org/10.1371/journal.pntd.0001505 |
| [42] | Akoto C, Davies DE, Swindle EJ (2017) Mast cells are permissive for rhinovirus replication: potential implications for asthma exacerbations. Clin Exp Allergy 47: 351-360. https://doi.org/10.1111/cea.12879 |
| [43] | Huo C, Wu H, Xiao J, et al. (2019) Genomic and bioinformatic characterization of mouse mast cells (P815) upon different influenza a virus (H1N1, H5N1, and H7N2) infections. Front Genet 10: 595. https://doi.org/10.3389/fgene.2019.00595 |
| [44] | Portales-Cervantes L, Haidl ID, Lee PW, et al. (2017) Virus-infected human mast cells enhance natural killer cell functions. J Innate Immun 9: 94-108. https://doi.org/10.1159/000450576 |
| [45] | Brisse M, Ly H (2019) Comparative structure and function analysis of the RIG-I-like receptors: RIG-I and MDA5. Front Immunol 10: 1586. https://doi.org/10.3389/fimmu.2019.01586 |
| [46] | St John AL, Rathore APS, Yap H, et al. (2011) Immune surveillance by mast cells during dengue infection promotes natural killer (NK) and NKT-cell recruitment and viral clearance. P Natl Acad Sci USA 108: 9190-9195. https://doi.org/10.1073/pnas.1105079108 |
| [47] | Graham AC, Temple RM, Obar JJ (2015) Mast cells and influenza a virus: association with allergic responses and beyond. Front Immunol 6: 238. https://doi.org/10.3389/fimmu.2015.00238 |
| [48] | Dillon SR, Sprecher C, Hammond A, et al. (2004) Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol 5: 752-760. https://doi.org/10.1038/ni1084 |
| [49] | Zhang Q, Putheti P, Zhou Q, et al. (2008) Structures and biological functions of IL-31 and IL-31 receptors. Cytokine Growth Factor Rev 19: 347-356. https://doi.org/10.1016/j.cytogfr.2008.08.003 |
| [50] | Gangemi S, Franchina T, Minciullo PL, et al. (2013) IL-33/IL-31 axis: a new pathological mechanisms for EGFR tyrosine kinase inhibitors-associated skin toxicity. J Cell Biochem 114: 2673-2676. https://doi.org/10.1002/jcb.24614 |
| [51] | Guarneri F, Minciullo PL, Mannucci C, et al. (2015) IL-31 and IL-33 circulating levels in allergic contact dermatitis. Eur Ann Allergy Clin Immunol 47: 156-158. |
| [52] | Bonanno A, Gangemi S, La Grutta S, et al. (2014) 25-Hydroxyvitamin D, IL-31, and IL-33 in children with allergic disease of the airways. Mediat Inflamm 2014: 520241. https://doi.org/10.1155/2014/520241 |
| [53] | Angulo EL, McKernan EM, Fichtinger PS, et al. (2019) Comparison of IL-33 and IL-5 family mediated activation of human eosinophils. PLoS One 14: e0217807. https://doi.org/10.1371/journal.pone.0217807 |
| [54] | Stott B, Lavender P, Lehmann S, et al. (2013) Human IL-31 is induced by IL-4 and promotes Th2-driven inflammation. J Allergy Clin Immun 132: 446-454. https://doi.org/10.1016/j.jaci.2013.03.050 |
| [55] | Lai T, Wu D, Li W, et al. (2016) Interleukin-31 expression and relation to disease severity in human asthma. Sci Rep 6: 22835. https://doi.org/10.1038/srep22835 |
| [56] | Vocca L, Di Sano C, Uasuf CG, et al. (2015) IL-33/ST2 axis controls Th2/IL-31 and Th17 immune response in allergic airway diseases. Immunobiology 220: 954-963. https://doi.org/10.1016/j.imbio.2015.02.005 |
| [57] | Musolino C, Allegra A, Mannucci C, et al. (2015) Possible role of interleukin-31/33 axis in imatinib mesylate-associated skin toxicity. Turk J Haematoly 32: 168-171. https://doi.org/10.4274/Tjh.2014.0021 |
| [58] | Nygaard U, Hvid M, Johansen C, et al. (2016) TSLP, IL-31, IL-33 and sST2 are new biomarkers in endophenotypic profiling of adult and childhood atopic dermatitis. J Eur Acad Dermatol 30: 1930-1938. https://doi.org/10.1111/jdv.13679 |
| [59] | Bruhs A, Proksch E, Schwarz T, et al. (2018) Disruption of the epidermal barrier induces regulatory T cells via IL-33 in mice. J Invest Dermatol 138: 570-579. https://doi.org/10.1016/j.jid.2017.09.032 |
| [60] | Wang Z, Yi T, Long M, et al. (2018) Involvement of the negative feedback of IL-33 signaling in the anti-inflammatory effect of electro-acupuncture on allergic contact dermatitis via targeting MicroRNA-155 in mast cells. Inflammation 41: 859-869. https://doi.org/10.1007/s10753-018-0740-8 |
| [61] | Liu B, Tai Y, Achanta S, et al. (2016) IL-33/ST2 signaling excites sensory neurons and mediates itch response in a mouse model of poison ivy contact allergy. P Natl Acad Sci USA 113: E7572-E7579. https://doi.org/10.1073/pnas.1606608113 |
| [62] | Murdaca G, Greco M, Tonacci A, et al. (2019) IL-33/IL-31 axis in immune-mediated and allergic diseases. Int J Mol Sci 20: 5856. https://doi.org/10.3390/ijms20235856 |
| [63] | Murdaca G, Allegra A, Tonacci A, et al. (2022) Mast cells and vitamin D status: A clinical and biological link in the onset of allergy and bone diseases. Biomedicines 10: 1877. https://doi.org/10.3390/biomedicines10081877 |
| [64] | Heine G, Niesner U, Chang HD, et al. (2008) 1,25-dihydroxyvitamin D(3) promotes IL-10 production in human B cells. Eur J Immunol 38: 2210-2218. https://doi.org/10.1002/eji.200838216 |
| [65] | Drozdenko G, Scheel T, Heine G, et al. (2014) Impaired T cell activation and cytokine production by calcitriol-primed human B cells. Clin Exp Immunol 178: 364-372. https://doi.org/10.1111/cei.12406 |
| [66] | Liu Z, Li X, Qiu S, et al. (2017) Vitamin D contributes to mast cell stabilization. Allergy 72: 1184-1192. https://doi.org/10.1111/all.13110 |
| [67] | Biggs L, Yu C, Fedoric B, et al. (2010) Evidence that vitamin D(3) promotes mast cell-dependent reduction of chronic UVB-induced skin pathology in mice. J Exp Med 207: 455-463. https://doi.org/10.1084/jem.20091725 |
| [68] | Asero R, Ferrucci S, Casazza G, et al. (2019) Total IgE and atopic status in patients with severe chronic spontaneous urticaria unresponsive to omalizumab treatment. Allergy 74: 1561-1563. https://doi.org/10.1111/all.13754 |
| [69] | Lakin E, Church MK, Maurer M, et al. (2019) On the lipophilic nature of autoreactive IgE in chronic spontaneous urticaria. Theranostics 9: 829-836. https://doi.org/10.7150/thno.29902 |
| [70] | Redegeld FA, Yu Y, Kumari S, et al. (2018) Non-IgE mediated mast cell activation. Immunol Rev 282: 87-113. https://doi.org/10.1111/imr.12629 |
| [71] | Bakdash G, van Capel TM, Mason LM, et al. (2014) Vitamin D3 metabolite calcidiol primes human dendritic cells to promote the development of immunomodulatory IL-10-producing T cells. Vaccine 32: 6294-6302. https://doi.org/10.1016/j.vaccine.2014.08.075 |
| [72] | Almerighi C, Sinistro A, Cavazza A, et al. (2009) 1α,25-dihydroxyvitamin D3 inhibits CD40L-induced pro-inflammatory and immunomodulatory activity in human monocytes. Cytokine 45: 190-197. https://doi.org/10.1016/j.cyto.2008.12.009 |
| [73] | Ly NP, Litonjua A, Gold DR, et al. (2011) Gut microbiota, probiotics, and vitamin D: interrelated exposures influencing allergy, asthma, and obesity?. J Allergy Clin Immun 127: 1087-1094. https://doi.org/10.1016/j.jaci.2011.02.015 |
| [74] | Suvorov A (2013) Gut microbiota, probiotics, and human health. Biosci Microb Food H 32: 81-91. https://doi.org/10.12938/bmfh.32.81 |
| [75] | Traina G (2021) The role of mast cells in the gut and brain. J Integr Neurosci 20: 185-196. https://doi.org/10.31083/j.jin.2021.01.313 |
| [76] | Conte C, Sichetti M, Traina G (2020) Gut–brain axis: focus on neurodegeneration and mast cells. Appl Sci 10: 1828. https://doi.org/10.3390/app10051828 |
| [77] | Lynn DJ, Benson SC, Lynn MA, et al. (2022) Modulation of immune responses to vaccination by the microbiota: implications and potential mechanisms. Nat Rev Immunol 22: 33-46. https://doi.org/10.1038/s41577-021-00554-7 |
| [78] | Hu Y, Jin Y, Han D, et al. (2012) Mast cell-induced lung injury in mice infected with H5N1 influenza virus. J Virol 86: 3347-3356. https://doi.org/10.1128/JVI.06053-11 |
| [79] | Zarnegar B, Westin A, Evangelidou S, et al. (2018) Innate immunity induces the accumulation of lung mast cells during influenza infection. Front Immunol 9: 2288. https://doi.org/10.3389/fimmu.2018.02288 |
| [80] | Liu B, Meng D, Wei T, et al. (2014) Apoptosis and pro-inflammatory cytokine response of mast cells induced by influenza A viruses. PLoS One 9: e100109. https://doi.org/10.1371/journal.pone.0100109 |
| [81] | Wu H, Zhang S, Huo C, et al. (2019) iTRAQ-based proteomic and bioinformatic characterization of human mast cells upon infection by the influenza A virus strains H1N1 and H5N1. FEBS Lett 593: 2612-2627. https://doi.org/10.1002/1873-3468.13523 |
| [82] | Ng K, Raheem J, St Laurent CD, et al. (2019) Responses of human mast cells and epithelial cells following exposure to influenza A virus. Antivir Res 171: 104566. https://doi.org/10.1016/j.antiviral.2019.104566 |
| [83] | Kulka M, Alexopoulou L, Flavell RA, et al. (2004) Activation of mast cells by double-stranded RNA: evidence for activation through Toll-like receptor 3. J Allergy Clin Immun 114: 174-182. https://doi.org/10.1016/j.jaci.2004.03.049 |
| [84] | Pelaia G, Vatrella A, Gallelli L, et al. (2006) Respiratory infections and asthma. Resp Med 100: 775-784. https://doi.org/10.1016/j.rmed.2005.08.025 |
| [85] | Al-Afif A, Alyazidi R, Oldford SA, et al. (2015) Respiratory syncytial virus infection of primary human mast cells induces the selective production of type I interferons, CXCL10, and CCL4. J Allergy Clin Immun 136: 1346-1354. https://doi.org/10.1016/j.jaci.2015.01.042 |
| [86] | Reeves SR, Barrow KA, Rich LM, et al. (2020) Respiratory syncytial virus infection of human lung fibroblasts induces a hyaluronan-enriched extracellular matrix that binds mast cells and enhances expression of mast cell proteases. Front Immunol 10: 3159. https://doi.org/10.3389/fimmu.2019.03159 |
| [87] | Hosoda M, Yamaya M, Suzuki T, et al. (2002) Effects of rhinovirus infection on histamine and cytokine production by cell lines from human mast cells and basophils. J Immunol 169: 1482-1491. https://doi.org/10.4049/jimmunol.169.3.1482 |
| [88] | Liu H, Tan J, Liu J, et al. (2020) Altered mast cell activity in response to rhinovirus infection provides novel insight into asthma. J Asthma 57: 459-467. https://doi.org/10.1080/02770903.2019.1585870 |
| [89] | Kritas SK, Ronconi G, Caraffa AL, et al. (2020) Mast cells contribute to coronavirus-induced inflammation: new anti-inflammatory strategy. J Biol Regul Homeost Agents 34: 9-14. |
| [90] | Theoharides TC, Tsilioni I, Ren H (2019) Recent advances in our understanding of mast cell activation–or should it be mast cell mediator disorders?. Expert Rev Clin Immunol 15: 639-656. https://doi.org/10.1080/1744666X.2019.1596800 |
| [91] | Theoharides TC (2021) Potential association of mast cells with coronavirus disease 2019. Ann Allerg Asthma Im 126: 217-218. https://doi.org/10.1016/j.anai.2020.11.003 |
| [92] | Kempuraj D, Selvakumar GP, Ahmed ME, et al. (2020) COVID-19, mast cells, cytokine storm, psychological stress, and neuroinflammation. Neuroscientist 26: 402-414. https://doi.org/10.1177/1073858420941476 |
| [93] | Junior JSM, Miggiolaro AFRDS, Nagashima S, et al. (2020) Mast cells in alveolar septa of COVID-19 patients: a pathogenic pathway that may link interstitial edema to immunothrombosis. Front Immunol 11: 574862. https://doi.org/10.3389/fimmu.2020.574862 |
| [94] | Ricke DO, Gherlone N, Fremont-Smith P, et al. (2020) Kawasaki disease, multisystem inflammatory syndrome in children: antibody-induced mast cell activation hypothesis. J Pediatrics Pediatr Med 4: 1-7. https://doi.org/10.29245/2578-2940/2020/2.1157 |
| [95] | Afrin LB, Weinstock LB, Molderings GJ (2020) Covid-19 hyperinflammation and post-Covid-19 illness may be rooted in mast cell activation syndrome. Int J Infect Dis 100: 327-332. https://doi.org/10.1016/j.ijid.2020.09.016 |
| [96] | Weinstock LB, Brook JB, Walters AS, et al. (2021) Mast cell activation symptoms are prevalent in Long-COVID. Int J Infect Dis 112: 217-226. https://doi.org/10.1016/j.ijid.2021.09.043 |
| [97] | Nagaraja V, Matucci-Cerinic M, Furst DE, et al. (2020) Current and future outlook on disease modification and defining low disease activity in systemic sclerosis. Arthritis Rheumatol 72: 1049-1058. https://doi.org/10.1002/art.41246 |
| [98] | Arnold J, Winthrop K, Emery P (2021) COVID-19 vaccination and antirheumatic therapy. Rheumatology 60: 3496-3502. https://doi.org/10.1093/rheumatology/keab223 |
| [99] | Creech CB, Walker SC, Samuels RJ (2021) SARS-CoV-2 vaccines. JAMA 325: 1318. https://doi.org/10.1001/jama.2021.3199 |
| [100] | Hazlewood GS, Pardo JP, Barnabe C, et al. (2021) Canadian rheumatology association recommendation for the use of COVID-19 vaccination for patients with autoimmune rheumatic diseases. J Rheumatol 48: 1330-1339. https://doi.org/10.3899/jrheum.210288 |
| [101] | European Medicines Agency.Comirnaty and Spikevax: EMA Recommendations on Extra Doses Boosters. European Medicines Agency (2021) . Available from: https://www.ema.europa.eu/en/news/comirnaty-spikevax-ema-recommendations-extra-doses-boosters. |
| [102] | Elhai M, Avouac J, Walker U, et al. (2016) A gender gap in primary and secondary heart dysfunctions in systemic sclerosis: A EUSTAR prospective study. Ann Rheum Dis 75: 163-169. https://doi.org/10.1136/annrheumdis-2014-206386 |
| [103] | Khedoe P, Marges E, Hiemstra P, et al. (2020) Interstitial lung disease in patients with systemic sclerosis: Toward personalized-medicine-based prediction and drug screening models of systemic sclerosis-related interstitial lung disease (SSc-ILD). Front Immunol 11: 19090. https://doi.org/10.3389/fimmu.2020.01990 |
| [104] | Alba MA, Velasco C, Simeón CP, et al. (2014) Early-versus late-onset systemic sclerosis: differences in clinical presentation and outcome in 1037 patients. Medicine 93: 73-81. https://doi.org/10.1097/MD.0000000000000018 |
| [105] | Murdaca G, Noberasco G, Olobardi D, et al. (2021) Current take on systemic sclerosis patients' vaccination recommendations. Vaccines 9: 1426. https://doi.org/10.3390/vaccines9121426 |
| [106] | Weinreich DM, Sivapalasingam S, Norton T, et al. (2021) REGN-COV2, a neutralizing antibody cocktail, in outpatients with COVID-19. N Engl J Med 384: 238-251. https://doi.org/10.1056/NEJMoa2035002 |
| [107] | Iketani S, Liu L, Guo Y, et al. (2022) Antibody evasion properties of the SARS-CoV-2 omicron sublineages. Nature 604: 553-556. https://doi.org/10.1038/s41586-022-04594-4 |
| [108] | Hirsch C, Park YS, Piechotta V, et al. (2022) SARS-CoV-2-neutralising monoclonal antibodies to prevent COVID-19. Cochrane Datebase Syst Rev 2021: CD014945. https://doi.org/10.1002/14651858.CD014945 |
| [109] | Gordon JK, Showalter K, Wu Y, et al. (2022) Systemic sclerosis and COVID-19 vaccines: a SPIN cohort study. Lancet Rheumatol 4: e243-e246. https://doi.org/10.1016/S2665-9913(21)00416-1 |
| [110] | Sampaio-Barros PD, Medeiros-Ribeiro AC, Luppino-Assad AP, et al. (2022) SARS-CoV-2 vaccine in patients with systemic sclerosis: impact of disease subtype and therapy. Rheumatology 61: SI169-SI174. https://doi.org/10.1093/rheumatology/keab886 |
| [111] | Aikawa NE, Kupa LDVK, Medeiros-Ribeiro AC, et al. (2022) Increment of immunogenicity after third dose of a homologous inactivated SARS-CoV-2 vaccine in a large population of patients with autoimmune rheumatic diseases. Ann Rheum Dis 81: 1036-1043. https://doi.org/10.1136/annrheumdis-2021-222096 |
| [112] | Ferri C, Ursini F, Gragnani L, et al. (2021) Impaired immunogenicity to COVID-19 vaccines in autoimmune systemic diseases. High prevalence of non-response in different patients' subgroups. J Autoimmun 125: 102744. https://doi.org/10.1016/j.jaut.2021.102744 |
| [113] | Braun-Moscovici Y, Kaplan M, Braun M, et al. (2021) Disease activity and humoral response in patients with inflammatory rheumatic diseases after two doses of the Pfizer mRNA vaccine against SARS-CoV-2. Ann Rheum Dis 80: 1317-1321. https://doi.org/10.1136/annrheumdis-2021-220503 |
| [114] | Taghinezhad-S S, Mohseni AH, Bermúdez-Humarán LG, et al. (2021) Probiotic-based vaccines may provide effective protection against COVID-19 acute respiratory disease. Vaccines 9: 466. https://doi.org/10.3390/vaccines9050466 |
| [115] | Suvorov A, Gupalova T, Desheva Y, et al. (2021) Construction of the enterococcal strain expressing immunogenic fragment of SARS-Cov-2 virus. Front Pharmacol 12: 807256-807256. https://doi.org/10.3389/fphar.2021.807256 |
| [116] | Ubol S, Halstead SB (2010) How innate immune mechanisms contribute to antibody-enhanced viral infections. Clin Vaccine Immunol 17: 1829-1835. https://doi.org/10.1128/CVI.00316-10 |
| [117] | Taylor A, Foo SS, Bruzzone R, et al. (2015) Fc receptors in antibody-dependent enhancement of viral infections. Immunol Rev 268: 340-364. https://doi.org/10.1111/imr.12367 |
| [118] | Cardosa MJ, Porterfield JS, Gordon S (1983) Complement receptor mediates enhanced flavivirus replication in macrophages. J Exp Med 158: 258-263. https://doi.org/10.1084/jem.158.1.258 |
| [119] | Halstead SB, O'rourke EJ (1977) Antibody-enhanced dengue virus infection in primate leukocytes. Nature 265: 739-741. https://doi.org/10.1038/265739a0 |
| [120] | Anderson R (2003) Manipulation of cell surface macromolecules by flaviviruses. Adv Virus Res 59: 229. https://doi.org/10.1016/S0065-3527(03)59007-8 |
| [121] | Winarski KL, Tang J, Klenow L, et al. (2019) Antibody-dependent enhancement of influenza disease promoted by increase in hemagglutinin stem flexibility and virus fusion kinetics. P Natl Acad Sci USA 116: 15194-15199. https://doi.org/10.1073/pnas.1821317116 |
| [122] | Wang SF, Tseng SP, Yen CH, et al. (2014) Antibody-dependent SARS coronavirus infection is mediated by antibodies against spike proteins. Biochem Bioph Res Co 451: 208-214. https://doi.org/10.1016/j.bbrc.2014.07.090 |
| [123] | Kam YW, Kien F, Roberts A, et al. (2007) Antibodies against trimeric S glycoprotein protect hamsters against SARS-CoV challenge despite their capacity to mediate FcγRII-dependent entry into B cells in vitro. Vaccine 25: 729-740. https://doi.org/10.1016/j.vaccine.2006.08.011 |
| [124] | Wan Y, Shang J, Sun S, et al. (2020) Molecular mechanism for antibody-dependent enhancement of coronavirus entry. J Virol 94: e02015-19. https://doi.org/10.1128/JVI.02015-19 |
| [125] | Castilow EM, Olson MR, Varga SM (2007) Understanding respiratory syncytial virus (RSV) vaccine-enhanced disease. Immunol Res 39: 225-239. https://doi.org/10.1007/s12026-007-0071-6 |
| [126] | Polack FP, Teng MN, Collins PL, et al. (2002) A role for immune complexes in enhanced respiratory syncytial virus disease. J Exp Med 196: 859-865. https://doi.org/10.1084/jem.20020781 |
| [127] | Dakhama A, Park JW, Taube C, et al. (2004) The role of virus-specific immunoglobulin E in airway hyperresponsiveness. Am J Resp Crit Care 170: 952-959. https://doi.org/10.1164/rccm.200311-1610OC |
| [128] | Koraka P, Murgue B, Deparis X, et al. (2003) Elevated levels of total and dengue virus-specific immunoglobulin E in patients with varying disease severity. J Med Virol 70: 91-98. https://doi.org/10.1002/jmv.10358 |
| [129] | McKenna DB, Neill WA, Norval M (2001) Herpes simplex virus-specific immune responses in subjects with frequent and infrequent orofacial recrudescences. Br J Dermatol 144: 459-464. https://doi.org/10.1046/j.1365-2133.2001.04068.x |
| [130] | Votava M, Bartosova D, Krchnakova A, et al. (1996) Diagnostic importance of heterophile antibodies and immunoglobulins IgA, IgE, IgM and low-avidity IgG against Epstein-Barr virus capsid antigen in children. Acta Virol 40: 99-101. |
| [131] | Welliver RC, Wong DT, Middleton E, et al. (1982) Role of parainfluenza virus-specific IgE in pathogenesis of croup and wheezing subsequent to infection. J Pediatr 101: 889-896. https://doi.org/10.1016/S0022-3476(82)80005-X |
| [132] | Boonnak K, Slike BM, Burgess TH, et al. (2008) Role of dendritic cells in antibody-dependent enhancement of dengue virus infection. J Virol 82: 3939-3951. https://doi.org/10.1128/JVI.02484-07 |
| [133] | Skowronski DM, De Serres G, Crowcroft NS, et al. (2010) Association between the 2008–09 seasonal influenza vaccine and pandemic H1N1 illness during spring–summer 2009: four observational studies from Canada. PLoS Med 7: e1000258. https://doi.org/10.1371/journal.pmed.1000258 |
| [134] | Monsalvo AC, Batalle JP, Lopez MF, et al. (2011) Severe pandemic 2009 H1N1 influenza disease due to pathogenic immune complexes. Nat Med 17: 195-199. https://doi.org/10.1038/nm.2262 |
| [135] | Guihot A, Luyt CE, Parrot A, et al. (2014) Low titers of serum antibodies inhibiting hemagglutination predict fatal fulminant influenza A (H1N1) 2009 infection. Am J Resp Crit Care 189: 1240-1249. https://doi.org/10.1164/rccm.201311-2071OC |
| [136] | Kilbourne ED, Smith C, Brett I, et al. (2002) The total influenza vaccine failure of 1947 revisited: major intrasubtypic antigenic change can explain failure of vaccine in a post-World War II epidemic. P Natl Acad Sci USA 99: 10748-10752. https://doi.org/10.1073/pnas.162366899 |
| [137] | Ferdinands JM, Fry AM, Reynolds S, et al. (2017) Intraseason waning of influenza vaccine protection: evidence from the US influenza vaccine effectiveness network, 2011–2012 through 2014–2015. Clin Infect Dis 64: 544-550. https://doi.org/10.1093/cid/ciw816 |
| [138] | Escalera-Zamudio M, Cobián-Güemes G, de los Dolores Soto-del M, et al. (2012) Characterization of an influenza A virus in Mexican swine that is related to the A/H1N1/2009 pandemic clade. Virology 433: 176-182. https://doi.org/10.1016/j.virol.2012.08.003 |
| [139] | Rajao DS, Sandbulte MR, Gauger PC, et al. (2016) Heterologous challenge in the presence of maternally-derived antibodies results in vaccine-associated enhanced respiratory disease in weaned piglets. Virology 491: 79-88. https://doi.org/10.1016/j.virol.2016.01.015 |
| [140] | Khurana S, Loving CL, Manischewitz J, et al. (2013) Vaccine-induced anti-HA2 antibodies promote virus fusion and enhance influenza virus respiratory disease. Sci Transl Med 5: 200ra114. https://doi.org/10.1126/scitranslmed.3006366 |
| [141] | Arvin AM, Fink K, Schmid MA, et al. (2020) A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 584: 353-363. https://doi.org/10.1038/s41586-020-2538-8 |
| [142] | Lee WS, Wheatley AK, Kent SJ, et al. (2020) Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies. Nat Microbiol 5: 1185-1191. https://doi.org/10.1038/s41564-020-00789-5 |
| [143] | Wen J, Cheng Y, Ling R, et al. (2020) Antibody-dependent enhancement of coronavirus. Int J Infect Dis 100: 483-489. https://doi.org/10.1016/j.ijid.2020.09.015 |
| [144] | Yip MS, Leung NHL, Cheung CY, et al. (2014) Antibody-dependent infection of human macrophages by severe acute respiratory syndrome coronavirus. Virol J 11: 1-11. https://doi.org/10.1186/1743-422X-11-82 |
| [145] | Bauer BS, Kerr ME, Sandmeyer LS, et al. (2013) Positive immunostaining for feline infectious peritonitis (FIP) in a Sphinx cat with cutaneous lesions and bilateral panuveitis. Vet Ophthalmol 16: 160-163. https://doi.org/10.1111/vop.12044 |
| [146] | Takano T, Kawakami C, Yamada S, et al. (2008) Antibody-dependent enhancement occurs upon re-infection with the identical serotype virus in feline infectious peritonitis virus infection. J Vet Med Sci 70: 1315-1321. https://doi.org/10.1292/jvms.70.1315 |
| [147] | Vennema H, Poland A, Foley J, et al. (1998) Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 243: 150-157. https://doi.org/10.1006/viro.1998.9045 |
| [148] | Harvima IT, Levi-Schaffer F, Draber P, et al. (2014) Molecular targets on mast cells and basophils for novel therapies. J Allergy Clin Immunol 134: 530-544. https://doi.org/10.1016/j.jaci.2014.03.007 |
| [149] | Fidan C, Aydoğdu A (2020) As a potential treatment of COVID-19: Montelukast. Med Hypotheses 142: 109828. https://doi.org/10.1016/j.mehy.2020.109828 |
| [150] | Malone RW, Tisdall P, Fremont-Smith P, et al. (2021) COVID-19: famotidine, histamine, mast cells, and mechanisms. Front Pharmacol 12: 633680. https://doi.org/10.3389/fphar.2021.633680 |
| [151] | Ennis M, Tiligada K (2021) Histamine receptors and COVID-19. Inflamm Res 70: 67-75. https://doi.org/10.1007/s00011-020-01422-1 |
| [152] | Han NR, Moon PD, Nam SY, et al. (2016) Inhibitory effects of atractylone on mast cell-mediated allergic reactions. Chem-Biol Interact 258: 59-68. https://doi.org/10.1016/j.cbi.2016.08.015 |
| [153] | Murphy-Schafer AR, Paust S (2021) Divergent mast cell responses modulate antiviral immunity during influenza virus infection. Front Cell Infect Microbiol 11: 580679. https://doi.org/10.3389/fcimb.2021.580679 |
| [154] | Shale M, Czub M, Kaplan GG, et al. (2010) Anti-tumor necrosis factor therapy and influenza: keeping it in perspective. Therap Adv Gastroenterol 3: 173-177. https://doi.org/10.1177/1756283X10366368 |
| [155] | Hoogeveen MJ, van Gorp EC, Hoogeveen EK (2020) Can pollen explain the seasonality of flu-like illnesses in the Netherlands?. Sci Total Environ 755: 143182. https://doi.org/10.1016/j.scitotenv.2020.143182 |
Figures(1)

Andrey Mamontov, Alexander Polevshchikov, Yulia Desheva. Mast cells in severe respiratory virus infections: insights for treatment and vaccine administration[J]. AIMS Allergy and Immunology, 2023, 7(1): 1-23. doi: 10.3934/Allergy.2023001







 DownLoad:
DownLoad: