Citation: Muriel Jourdan, Julia Pingel, Arnaud Buhot, Thierry Livache, Jean-François Constant. Surface plasmon resonance imaging of the conversion of clustered DNA lesions into double strand breaks by Fpg protein[J]. AIMS Materials Science, 2015, 2(4): 473-483. doi: 10.3934/matersci.2015.4.473

| [1] | Magnander K,Elmroth K(2012)Biological consequences of formation and repair of complex DNA damage.Cancer Lett 327: 90-96. |
| [2] | Eccles LJ, O'Neill P, Lomax ME (2011) Delayed repair of radiation induced clustered DNA damage: Friend or foe? MutatRes 711: 134-141. |
| [3] | Sage E, Harrison L (2011) Clustered DNA lesion repair in eukaryotes: Relevance to mutagenesis and cell survival.MutatRes 711: 123-133. |
| [4] | Okayasu R (2012) Repair of DNA damage induced by accelerated heavy ions-a mini review.IntJCancer 130: 991-1000. |
| [5] | Georgakilas AG, O'Neill P, Stewart RD (2013) Induction and repair of clustered DNA lesions: what do we know so far?RadiatRes 180: 100-109. |
| [6] | Pouget JP, Frelon S,Ravanat JL,et al. (2002) Formation of modified DNA bases in cells exposed either to gamma radiation or to high-LET particles.RadiatRes 157: 589-595. |
| [7] | Cadet J,Douki T,Ravanat JL(2008) Oxidatively generated damage to the guanine moiety of DNA: mechanic aspects and formation in cells.AccChemRes 41: 1075-1083. |
| [8] | Frelon S,Douki T,Ravanat JL,et al. (2000) High-performance liquid chromatography-tandem mass spectrometry measurement of radiation-induced base damage to isolated and cellular DNA.ChemResToxicol 13: 1002-1010. |
| [9] | Memisoglu A, Samson L(2000) Base excision repair in yeast and mammals.MutatRes 451: 39-51. |
| [10] | Wilson DM, Sofinowski TM and McNeill DR (2003) Repair mechanisms for oxidative DNA damage.FrontBiosci 8:963-981. |
| [11] | Shikazono N,Noguchi M,Fujii K,et al. (2009) The Yield, processing, and biological consequences of custered DNA camage induced by ionizing radiation.JRadiatRes 50: 27-36. |
| [12] |
Blaisdell JO, Wallace SS (2001) Abortive base-excision repair of radiation-induced clustered DNA lesions in Escherichia coli.ProcNatlAcadSciUSA 98: 7426-7430. doi: 10.1073/pnas.131077798

|
| [13] | D'Souza DI, Harrison L(2003) Repair of clustered uracil DNA damages in Escherichia coli.Nucleic Acids Res 31: 4573-4581. |
| [14] | Lomax ME,Cunniffe S,O'Neill P(2004) Efficiency of repair of an abasic site within DNA clustered damage sites by mammalian cell nuclear extracts.DNA Repair 43: 11017-11026. |
| [15] |
Pearson CG,Shikazono N,Tacker J,et al. (2004) Enhanced mutagenic potential of 8-oxo-7,8-dihydroguanine when present within a clustered DNA damage site.Nucleic Acids Res 32: 263-270. doi: 10.1093/nar/gkh150
|
| [16] | Budworth H,Dianova II,Podust VN,et al. (2002) Repair of clustered DNA lesions.JBiolChem 277: 21300-21305. |
| [17] | Tian K,McTigue M, de los Santos C(2002) Sorting the consequences of ionizing radiation: processing of 8-oxoguanine/abasic site lesions.DNA Repair 1: 1039-1049. |
| [18] |
Corne C, Fiche JB,Gasparutto D,et al. (2008) SPR imaging for label-free multiplexed analyses of DNA N-glycosylase interactions with damage DNA duplexes.Analyst 133: 1036-1045. doi: 10.1039/b801086b
|
| [19] |
David-Cordonnier MH, Laval J, O'Neill P (2001) Recognition and kinetics for excision of a base lesion within clustered DNA damage by the Escherichia coli proteins Fpg and Nth.Biochemistry 40: 5738-5746. doi: 10.1021/bi002605d
|
| [20] | David-Cordonnier MH, Laval J, O'Neill P(2000) Clustered DNA damage, influence on damage excision by XRS5 nuclear extracts and Escherichia coli Nth and Fpg proteins.JBiolChem 275: 11865-11873. |
| [21] | Harisson L, Hatahet Z, Wallace SS (1999) In vitro repair of synthetic ionizing radiation-induced multiply damaged DNA sites.JMolBiol 290: 667-684. |
| [22] | Zálešák J,Lourdin M,Krejčί L,et al. (2014) Structure and dynamics of DNA duplexes containing a cluster of mutagenic 8-oxoguanine and abasic site lesions.JMolBiol 426: 1524-1538. |
| [23] |
BhartiSK,Varshney U (2010) Analysis of the impact of a uracil DNA glycosylaseattenuated in AP-DNA binding in maintenance of the genomic integrity in Escherichia coli.Nucleic Acids Res 38: 2291-2301 doi: 10.1093/nar/gkp1210
|
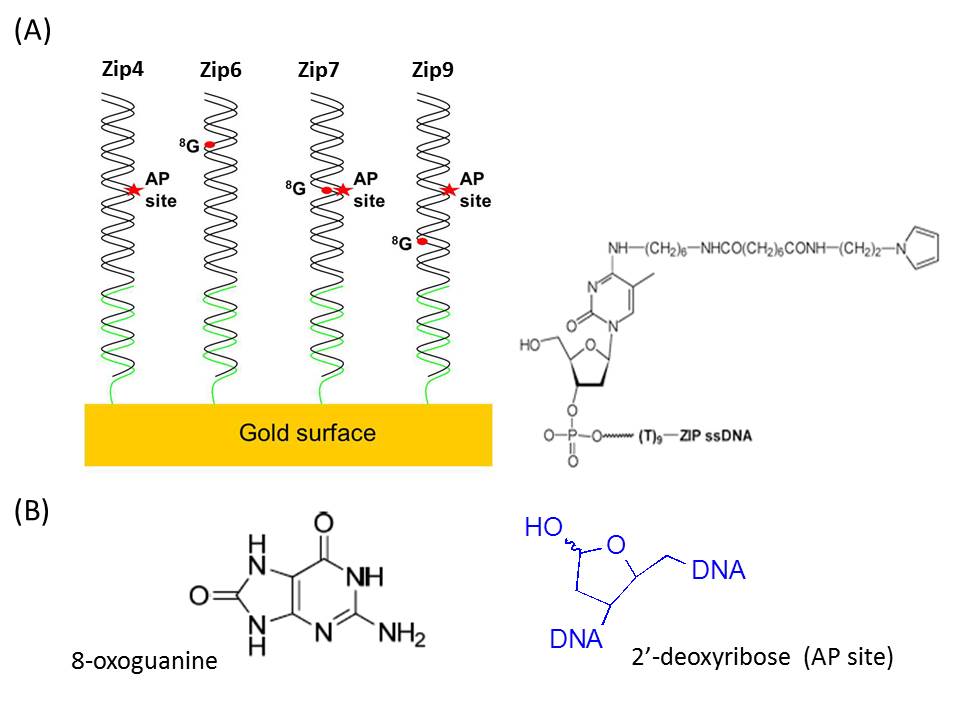
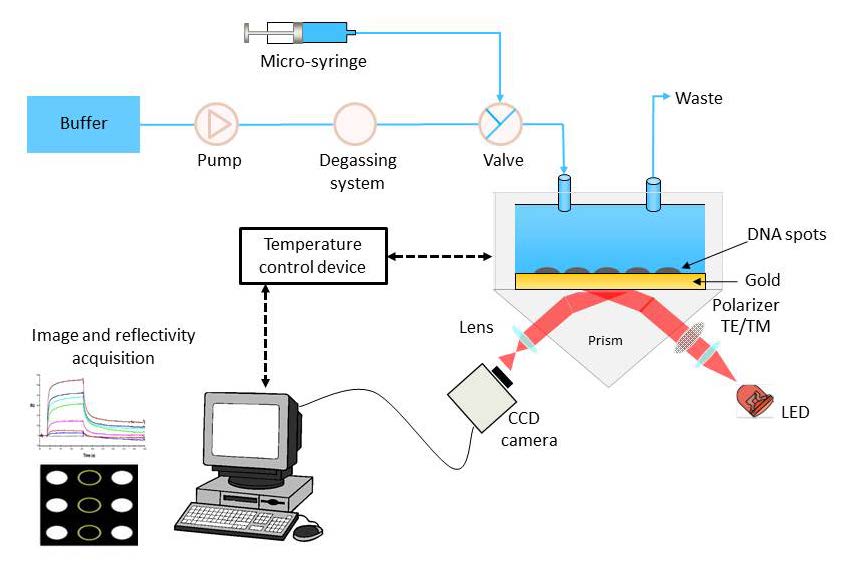


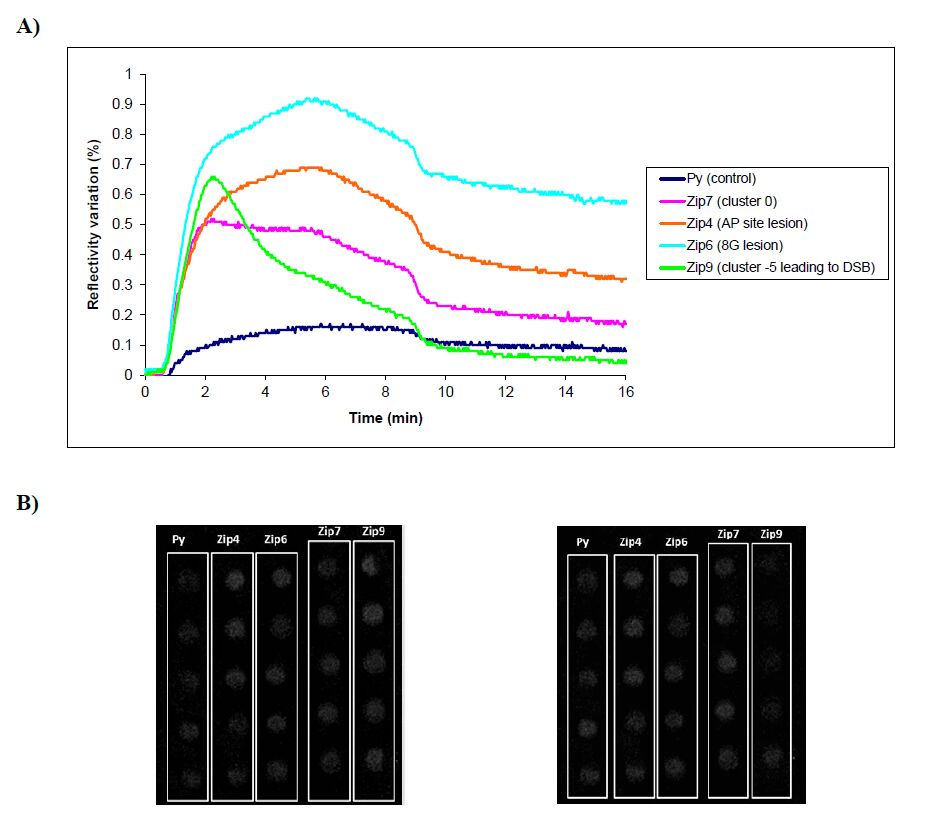
Figures(5) / Tables(2)

Muriel Jourdan, Julia Pingel, Arnaud Buhot, Thierry Livache, Jean-François Constant. Surface plasmon resonance imaging of the conversion of clustered DNA lesions into double strand breaks by Fpg protein[J]. AIMS Materials Science, 2015, 2(4): 473-483. doi: 10.3934/matersci.2015.4.473







 DownLoad:
DownLoad: