Citation: Daniel Sultanov, Nadezhda Gerasimova, Kseniya Kudryashova, Natalya Maluchenko, Elena Kotova, Marie-France Langelier, John Pascal, Mikhail Kirpichnikov, Alexey Feofanov, Vasily Studitsky. Unfolding of core nucleosomes by PARP-1 revealed by spFRET microscopy[J]. AIMS Genetics, 2017, 4(1): 21-31. doi: 10.3934/genet.2017.1.21

| [1] |
Beard BC, Wilson SH, Smerdon MJ (2003) Suppressed catalytic activity of base excision repair enzymes on rotationally positioned uracil in nucleosomes. Proc Natl Acad Sci U S A 100: 7465-7470. doi: 10.1073/pnas.1330328100

|
| [2] |
Caldecott KW (2007) Mammalian single-strand break repair: mechanisms and links with chromatin. DNA Repair (Amst) 6: 443-453. doi: 10.1016/j.dnarep.2006.10.006
|
| [3] |
De Vos M, Schreiber V, Dantzer F (2012) The diverse roles and clinical relevance of PARPs in DNA damage repair: current state of the art. Biochem Pharmacol 84: 137-146. doi: 10.1016/j.bcp.2012.03.018
|
| [4] |
Kraus WL (2008) Transcriptional control by PARP-1: chromatin modulation, enhancer-binding, coregulation, and insulation. Curr Opin Cell Biol 20: 294-302. doi: 10.1016/j.ceb.2008.03.006
|
| [5] |
Ali AA, Timinszky G, Arribas-Bosacoma R, et al. (2012) The zinc-finger domains of PARP1 cooperate to recognize DNA strand breaks. Nat Struct Mol Biol 19: 685-692. doi: 10.1038/nsmb.2335
|
| [6] |
Eustermann S, Wu WF, Langelier MF, et al. (2015) Structural Basis of Detection and Signaling of DNA Single-Strand Breaks by Human PARP-1. Mol Cell 60: 742-754. doi: 10.1016/j.molcel.2015.10.032
|
| [7] |
Langelier MF, Planck JL, Roy S, et al. (2012) Structural basis for DNA damage-dependent poly(ADP-ribosyl)ation by human PARP-1. Science 336: 728-732. doi: 10.1126/science.1216338
|
| [8] |
Martinez-Zamudio R, Ha HC (2012) Histone ADP-ribosylation facilitates gene transcription by directly remodeling nucleosomes. Mol Cell Biol 32: 2490-2502. doi: 10.1128/MCB.06667-11
|
| [9] |
Tanuma S, Kawashima K, Endo H (1985) Comparison of ADP-ribosylation of chromosomal proteins between intact and broken cells. Biochem Biophys Res Commun 127: 896-902. doi: 10.1016/S0006-291X(85)80028-0
|
| [10] |
Gunderson CC, Moore KN (2015) Olaparib: an oral PARP-1 and PARP-2 inhibitor with promising activity in ovarian cancer. Future Oncol 11: 747-757. doi: 10.2217/fon.14.313
|
| [11] |
Fong PC, Boss DS, Yap TA, et al. (2009) Inhibition of poly(ADP-ribose) polymerase in tumors from BRCA mutation carriers. N Engl J Med 361: 123-134. doi: 10.1056/NEJMoa0900212
|
| [12] |
Clark NJ, Kramer M, Muthurajan UM, et al. (2012) Alternative modes of binding of poly(ADP-ribose) polymerase 1 to free DNA and nucleosomes. J Biol Chem 287: 32430-32439. doi: 10.1074/jbc.M112.397067
|
| [13] |
Kudryashova KS, Chertkov OV, Nikitin DV, et al. (2015) Preparation of Mononucleosomal Templates for Analysis of Transcription with RNA Polymerase Using spFRET. Methods Mol Biol 1288: 395-412. doi: 10.1007/978-1-4939-2474-5_23
|
| [14] | Chertkov OV, Studitsky VM, Feofanov AV, et al. (2016) Change in conformation of linker DNA upon binding of histone Н1.5 to nucleosome: fluorescent microscopy of single complexes. Mosc Univ Biol Sci Bull 71: 108-113. |
| [15] |
Feofanov AV, Kudryashova KS, Chertkov OV, et al. (2015) Analysis of nucleosome transcription using single-particle FRET. Springer Proc Phys 164: 255-260. doi: 10.1007/978-3-319-16919-4_33
|
| [16] | Valieva ME, Armeev GA, Kudryashova KS, et al. (2016) Large-Scale ATP-Independent Nucleosome Unfolding by a Histone Chaperone. Nat Struct Mol Biol: In press. |
| [17] |
Langelier MF, Planck JL, Servent KM, et al. (2011) Purification of human PARP-1 and PARP-1 domains from Escherichia coli for structural and biochemical analysis. Methods Mol Biol 780: 209-226. doi: 10.1007/978-1-61779-270-0_13
|
| [18] |
Kulaeva OI, Gaykalova DA, Pestov NA, et al. (2009) Mechanism of chromatin remodeling and recovery during passage of RNA polymerase II. Nat Struct Mol Biol 16: 1272-1278. doi: 10.1038/nsmb.1689
|
| [19] |
Kireeva ML, Walter W, Tchernajenko V, et al. (2002) Nucleosome remodeling induced by RNA polymerase II: loss of the H2A/H2B dimer during transcription. Mol Cell 9: 541-552. doi: 10.1016/S1097-2765(02)00472-0
|
| [20] |
Studitsky VM, Clark DJ, Felsenfeld G (1995) Overcoming a nucleosomal barrier to transcription. Cell 83: 19-27. doi: 10.1016/0092-8674(95)90230-9
|
| [21] |
Vasudevan D, Chua EY, Davey CA (2010) Crystal structures of nucleosome core particles containing the '601' strong positioning sequence. J Mol Biol 403: 1-10. doi: 10.1016/j.jmb.2010.08.039
|
| [22] |
Thastrom A, Lowary PT, Widlund HR, et al. (1999) Sequence motifs and free energies of selected natural and non-natural nucleosome positioning DNA sequences. J Mol Biol 288: 213-229. doi: 10.1006/jmbi.1999.2686
|
| [23] |
Muthurajan UM, Hepler MR, Hieb AR, et al. (2014) Automodification switches PARP-1 function from chromatin architectural protein to histone chaperone. Proc Natl Acad Sci U S A 111: 12752-12757. doi: 10.1073/pnas.1405005111
|
| [24] | Zahradka P, Ebisuzaki K (1982) A shuttle mechanism for DNA-protein interactions. The regulation of poly(ADP-ribose) polymerase. Eur J Biochem 127: 579-585. |
| [25] | Ogata N, Ueda K, Kawaichi M, et al. (1981) Poly(ADP-ribose) synthetase, a main acceptor of poly(ADP-ribose) in isolated nuclei. J Biol Chem 256: 4135-4137. |
| [26] | Steffen JD, McCauley MM, Pascal JM (2016) Fluorescent sensors of PARP-1 structural dynamics and allosteric regulation in response to DNA damage. Nucleic Acids Res: gkw710. |
| [27] |
Dawicki-McKenna JM, Langelier MF, DeNizio JE, et al. (2015) PARP-1 Activation Requires Local Unfolding of an Autoinhibitory Domain. Mol Cell 60: 755-768. doi: 10.1016/j.molcel.2015.10.013
|
| [28] |
Lee JY, Lee TH (2012) Effects of DNA methylation on the structure of nucleosomes. J Am Chem Soc 134: 173-175. doi: 10.1021/ja210273w
|
| [29] |
Lee JY, Wei S, Lee TH (2011) Effects of histone acetylation by Piccolo NuA4 on the structure of a nucleosome and the interactions between two nucleosomes. J Biol Chem 286: 11099-11109. doi: 10.1074/jbc.M110.192047
|
| [30] | Chang HW, Shaytan AK, Hsieh FK, et al. (2013) Structural Analysis of the Key Intermediate Formed during Transcription through a Nucleosome. Trends Cell Mol Biol 8: 13-23. |
| [31] |
Pestov NA, Gerasimova NS, Kulaeva OI, et al. (2015) Structure of transcribed chromatin is a sensor of DNA damage. Sci Adv 1: e1500021. doi: 10.1126/sciadv.1500021
|
| [32] |
Gaykalova DA, Kulaeva OI, Volokh O, et al. (2015) Structural analysis of nucleosomal barrier to transcription. Proc Natl Acad Sci U S A 112: E5787-5795. doi: 10.1073/pnas.1508371112
|
| [33] |
Zaret KS, Carroll JS (2011) Pioneer transcription factors: establishing competence for gene expression. Genes Dev 25: 2227-2241. doi: 10.1101/gad.176826.111
|
| [34] |
Bohm V, Hieb AR, Andrews AJ, et al. (2011) Nucleosome accessibility governed by the dimer/tetramer interface. Nucleic Acids Res 39: 3093-3102. doi: 10.1093/nar/gkq1279
|
| [35] |
Tomschik M, Zheng H, van Holde K, et al. (2005) Fast, long-range, reversible conformational fluctuations in nucleosomes revealed by single-pair fluorescence resonance energy transfer. Proc Natl Acad Sci U S A 102: 3278-3283. doi: 10.1073/pnas.0500189102
|
| [36] |
Zlatanova J, Bishop TC, Victor JM, et al. (2009) The nucleosome family: dynamic and growing. Structure 17: 160-171. doi: 10.1016/j.str.2008.12.016
|
| [37] | Xu Y, Ayrapetov MK, Xu C, et al. (2012) Histone H2A.Z controls a critical chromatin remodeling step required for DNA double-strand break repair. Mol Cell 48: 723-733. |
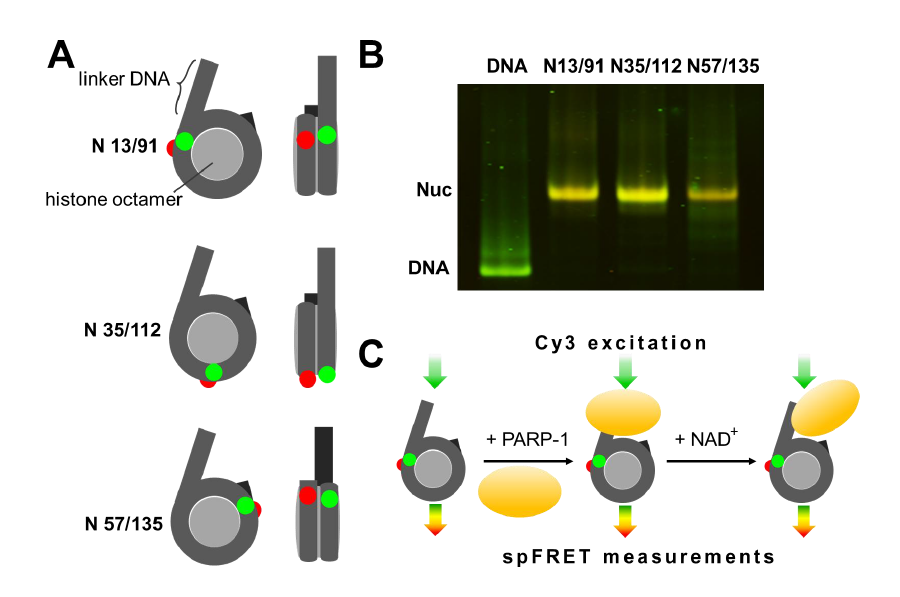
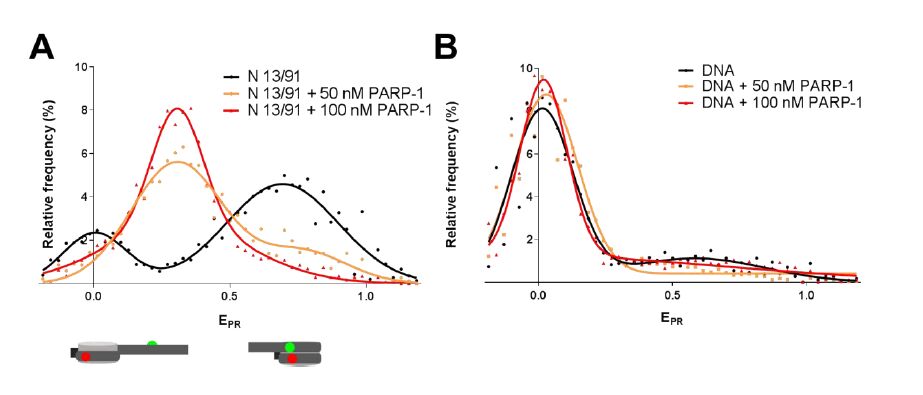
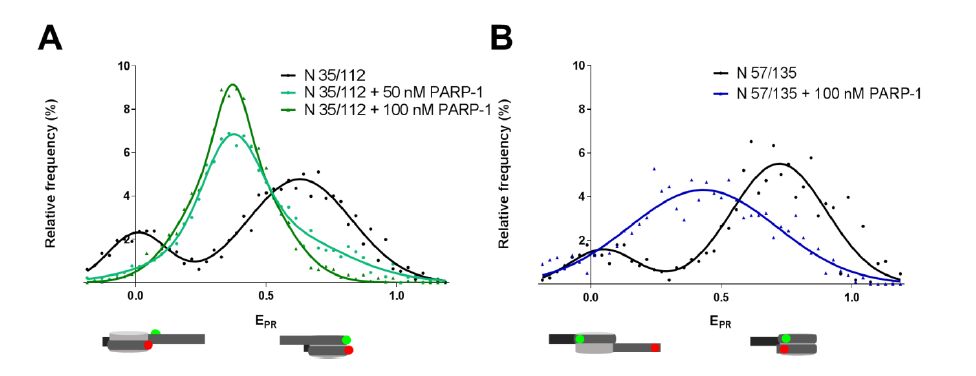
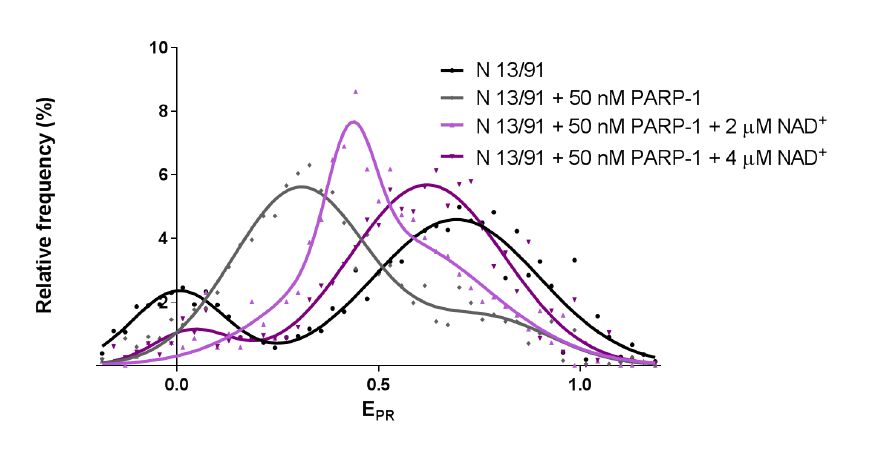
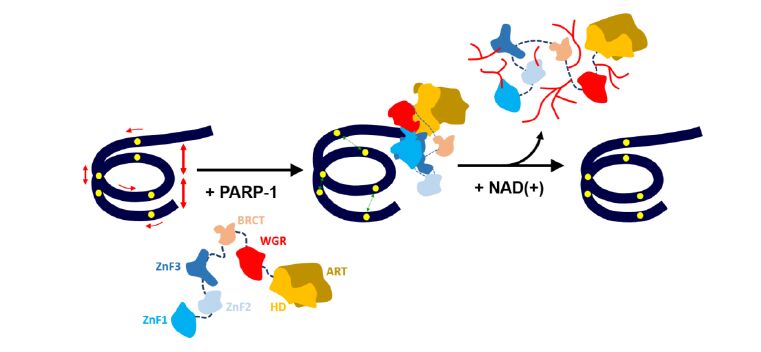
Figures(5)

Daniel Sultanov, Nadezhda Gerasimova, Kseniya Kudryashova, Natalya Maluchenko, Elena Kotova, Marie-France Langelier, John Pascal, Mikhail Kirpichnikov, Alexey Feofanov, Vasily Studitsky. Unfolding of core nucleosomes by PARP-1 revealed by spFRET microscopy[J]. AIMS Genetics, 2017, 4(1): 21-31. doi: 10.3934/genet.2017.1.21







 DownLoad:
DownLoad: