Non-consumptive effects such as fear of depredation, can strongly influence predator-prey dynamics. There are several ecological and social motivations for these effects in competitive systems as well. In this work we consider the classic two species ODE and PDE Lotka-Volterra competition models, where one of the competitors is "fearful" of the other. We find that the presence of fear can have several interesting dynamical effects on the classical competitive scenarios. Notably, for fear levels in certain regimes, we show novel bi-stability dynamics. Furthermore, in the spatially explicit setting, the effects of several spatially heterogeneous fear functions are investigated. In particular, we show that under certain integral restrictions on the fear function, a weak competition type situation can change to competitive exclusion. Applications of these results to ecological as well as sociopolitical settings are discussed, that connect to the "landscape of fear" (LOF) concept in ecology.
Citation: Vaibhava Srivastava, Eric M. Takyi, Rana D. Parshad. The effect of 'fear' on two species competition[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 8814-8855. doi: 10.3934/mbe.2023388
Non-consumptive effects such as fear of depredation, can strongly influence predator-prey dynamics. There are several ecological and social motivations for these effects in competitive systems as well. In this work we consider the classic two species ODE and PDE Lotka-Volterra competition models, where one of the competitors is "fearful" of the other. We find that the presence of fear can have several interesting dynamical effects on the classical competitive scenarios. Notably, for fear levels in certain regimes, we show novel bi-stability dynamics. Furthermore, in the spatially explicit setting, the effects of several spatially heterogeneous fear functions are investigated. In particular, we show that under certain integral restrictions on the fear function, a weak competition type situation can change to competitive exclusion. Applications of these results to ecological as well as sociopolitical settings are discussed, that connect to the "landscape of fear" (LOF) concept in ecology.

| [1] | M. W. Dictionary, Merriam-webster, Available from: https://www.merriam-webster.com/. |
| [2] |
H. S. Bracha, Freeze, flight, fight, fright, faint: Adaptationist perspectives on the acute stress response spectrum, CNS Spectrums, 9 (2004), 679–685. https://doi.org/10.1017/S1092852900001954 doi: 10.1017/S1092852900001954

|
| [3] |
W. Cresswell, Predation in bird populations, J. Ornithol., 152 (2011), 251–263. https://doi.org/10.1007/s10336-010-0638-1 doi: 10.1007/s10336-010-0638-1
|
| [4] |
M. J. Sheriff, S. D. Peacor, D. Hawlena, M. Thaker, Non-consumptive predator effects on prey population size: a dearth of evidence, J. Animal Ecol., 89 (2020), 1302–1316. https://doi.org/10.1111/1365-2656.13213 doi: 10.1111/1365-2656.13213
|
| [5] |
S. D. Peacor, B. L. Peckarsky, G. C. Trussell, J. R. Vonesh, Costs of predator-induced phenotypic plasticity: a graphical model for predicting the contribution of nonconsumptive and consumptive effects of predators on prey, Oecologia, 171 (2013), 1–10. https://doi.org/10.1007/s00442-012-2394-9 doi: 10.1007/s00442-012-2394-9
|
| [6] |
B. L. Peckarsky, P. A. Abrams, D. I. Bolnick, L. M. Dill, J. H. Grabowski, B. Luttbeg, et al., Revisiting the classics: considering nonconsumptive effects in textbook examples of predator–prey interactions, Ecology, 89 (2008), 2416–2425. https://doi.org/10.1890/07-1131.1 doi: 10.1890/07-1131.1
|
| [7] |
J. S. Brown, J. W. Laundré, M. Gurung, The ecology of fear: optimal foraging, game theory, and trophic interactions, J. Mammal., 80 (1999), 385–399. https://doi.org/10.2307/1383287 doi: 10.2307/1383287
|
| [8] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator–prey interactions, J. Math. Biol., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [9] |
P. A. Abrams, The evolution of predator-prey interactions: theory and evidence, Ann. Rev. Ecol. Syst., 31 (2000), 79–105. https://doi.org/10.1146/annurev.ecolsys.31.1.79 doi: 10.1146/annurev.ecolsys.31.1.79
|
| [10] |
P. Panday, N. Pal, S. Samanta, P. Tryjanowski, J. Chattopadhyay, Dynamics of a stage-structured predator-prey model: cost and benefit of fear-induced group defense, J. Theor. Biol., 528 (2021), 110846. https://doi.org/10.1016/j.jtbi.2021.110846 doi: 10.1016/j.jtbi.2021.110846
|
| [11] |
J. Wang, Y. Cai, S. Fu, W. Wang, The effect of the fear factor on the dynamics of a predator-prey model incorporating the prey refuge, Chaos Interdiscip. J. Nonlinear Sci., 29 (2019), 083109. https://doi.org/10.1063/1.5111121 doi: 10.1063/1.5111121
|
| [12] |
H. Zhang, Y. Cai, S. Fu, W. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 356 (2019), 328–337. https://doi.org/10.1016/j.amc.2019.03.034 doi: 10.1016/j.amc.2019.03.034
|
| [13] |
X. Wang, X. Zou, Modeling the fear effect in predator–prey interactions with adaptive avoidance of predators, Bull. Math. Biol., 79 (2017), 1325–1359. https://doi.org/10.1007/s11538-017-0287-0 doi: 10.1007/s11538-017-0287-0
|
| [14] |
P. Mikula, F. Morelli, R. K. Lučan, D. N. Jones, P. Tryjanowski, Bats as prey of diurnal birds: a global perspective, Mammal Rev., 46 (2016), 160–174. https://doi.org/10.1111/mam.12060 doi: 10.1111/mam.12060
|
| [15] |
R. M. Pringle, T. R. Kartzinel, T. M. Palmer, T. J. Thurman, K. Fox-Dobbs, C. C. Xu, et al., Predator-induced collapse of niche structure and species coexistence, Nature, 570 (2019), 58–64. https://doi.org/10.1038/s41586-019-1264-6 doi: 10.1038/s41586-019-1264-6
|
| [16] |
S. Pal, S. Majhi, S. Mandal, N. Pal, Role of fear in a predator–prey model with Beddington–Deangelis functional response, Z. Naturforsch. A, 74 (2019), 581–595. https://doi.org/10.1515/zna-2018-0449 doi: 10.1515/zna-2018-0449
|
| [17] |
M. Das, G. Samanta, A prey-predator fractional order model with fear effect and group defense, Int. J. Dyn. Control, 9 (2021), 334–349. https://doi.org/10.1080/21642583.2021.1907259 doi: 10.1080/21642583.2021.1907259
|
| [18] |
S. Pal, N. Pal, S. Samanta, J. Chattopadhyay, Effect of hunting cooperation and fear in a predator-prey model, Ecol. Complexity, 39 (2019), 100770. https://doi.org/10.1016/j.ecocom.2019.100770 doi: 10.1016/j.ecocom.2019.100770
|
| [19] | A. Yousef, A. A. Thirthar, A. L. Alaoui, P. Panja, T. Abdeljawad, The hunting cooperation of a predator under two prey's competition and fear-effect in the prey-predator fractional-order model, AIMS Math., 7 (2022), 5463–5479. |
| [20] |
P. Panday, N. Pal, S. Samanta, J. Chattopadhyay, Stability and bifurcation analysis of a three-species food chain model with fear, Int. J. Bifurcation Chaos, 28 (2018), 1850009. https://doi.org/10.1142/S0218127418500098 doi: 10.1142/S0218127418500098
|
| [21] |
P. Panday, N. Pal, S. Samanta, J. Chattopadhyay, A three species food chain model with fear induced trophic cascade, Int. J. Appl. Comput. Math., 5 (2019), 1–26. https://doi.org/10.1007/s40819-018-0585-8 doi: 10.1007/s40819-018-0585-8
|
| [22] |
H. Verma, K. Antwi-Fordjour, M. Hossain, N. Pal, R. D. Parshad, P. Mathur, A "double" fear effect in a tri-trophic food chain model, Eur. Phys. J. Plus, 136 (2021), 1–17. https://doi.org/10.1140/epjp/s13360-020-01001-7 doi: 10.1140/epjp/s13360-020-01001-7
|
| [23] |
A. Das, G. Samanta, Modeling the fear effect on a stochastic prey–predator system with additional food for the predator, J. Phys. A Math. Theor., 51 (2018), 465601. https://doi.org/10.1088/1751-8121/aae4c6 doi: 10.1088/1751-8121/aae4c6
|
| [24] |
S. Wu, J. Wang, J. Shi, Dynamics and pattern formation of a diffusive predator–prey model with predator-taxis, Math. Models Methods Appl. Sci., 28 (2018), 2275–2312. https://doi.org/10.1142/S0218202518400158 doi: 10.1142/S0218202518400158
|
| [25] |
X. Wang, X. Zou, Pattern formation of a predator-prey model with the cost of anti-predator behaviors, Math. Biosci. Eng., 15 (2018), 775. https://doi.org/10.1088/1742-2140/aaa076 doi: 10.1088/1742-2140/aaa076
|
| [26] |
D. Duan, B. Niu, J. Wei, Hopf-Hopf bifurcation and chaotic attractors in a delayed diffusive predator-prey model with fear effect, Chaos Solitons Fractals, 123 (2019), 206–216. https://doi.org/10.1016/j.chaos.2019.04.012 doi: 10.1016/j.chaos.2019.04.012
|
| [27] | Y. Lou, Some challenging mathematical problems in evolution of dispersal and population dynamics, in Tutorials in Mathematical Biosciences IV, (2008), 171–205. |
| [28] |
P. J. den Boer, The present status of the competitive exclusion principle, Trends Ecol. Evol., 1 (1986), 25–28. https://doi.org/10.1016/0169-5347(86)90064-9 doi: 10.1016/0169-5347(86)90064-9
|
| [29] |
P. Chesson, Mechanisms of maintenance of species diversity, Ann. Rev. Ecol. Syst., 31 (2000), 343–366. https://doi.org/10.1146/annurev.ecolsys.31.1.343 doi: 10.1146/annurev.ecolsys.31.1.343
|
| [30] |
A. Okubo, P. K. Maini, M. H. Williamson, J. D. Murray, On the spatial spread of the grey squirrel in Britain, Proc. R. Soc. London. B. Biol. Sci., 238 (1989), 113–125. https://doi.org/10.1098/rspb.1989.0070 doi: 10.1098/rspb.1989.0070
|
| [31] |
J. Sugie, R. Kohno, R. Miyazaki, On a predator-prey system of Holling type, Proc. Am. Math. Soc., 125 (1997), 2041–2050. https://doi.org/10.1090/S0002-9939-97-03901-4 doi: 10.1090/S0002-9939-97-03901-4
|
| [32] |
M. Arim, S. R. Abades, P. E. Neill, M. Lima, P. A. Marquet, Spread dynamics of invasive species, Proc. Nat. Acad. Sci., 103 (2006), 374–378. https://doi.org/10.1073/pnas.0504272102 doi: 10.1073/pnas.0504272102
|
| [33] |
G. A. Polis, C. A. Myers, R. D. Holt, The ecology and evolution of intraguild predation: potential competitors that eat each other, Ann. Rev. Ecol. Syst., 20 (1989), 297–330. https://doi.org/10.1146/annurev.es.20.110189.001501 doi: 10.1146/annurev.es.20.110189.001501
|
| [34] | X. Gu, K. Zhang, Z. Q. Zhang, Non-consumptive effects of intraguild predator Blattisocius dentriticus (Berlese) on the development and prey consumption of Neoseiulus cucumeris (Oudemans), Syst. Appl. Acarol., 27 (2022), 1475–1482. |
| [35] | R. Van Driesche, T. S. Bellows, Biological Control, Springer Science & Business Media, 2012. |
| [36] | L. L. Long, J. D. Wolfe, Review of the effects of barred owls on spotted owls, J. Wildlife Manage., 83 (2019), 1281–1296. |
| [37] |
N. J. Van Lanen, A. B. Franklin, K. P. Huyvaert, R. F. Reiser II, P. C. Carlson, Who hits and hoots at whom? Potential for interference competition between barred and northern spotted owls, Biol. Conserv., 144 (2011), 2194–2201. https://doi.org/10.1016/j.biocon.2011.05.011 doi: 10.1016/j.biocon.2011.05.011
|
| [38] | R. Gutiérrez, M. Cody, S. Courtney, D. Kennedy, Assessment of the potential threat of the northern barred owl, Sci. Eval. Status Northern Spotted Owl, 2004 (2004), 1–51. |
| [39] |
J. D. Wiens, R. G. Anthony, E. D. Forsman, Competitive interactions and resource partitioning between northern spotted owls and barred owls in western Oregon, Wildlife Monogr., 185 (2014), 1–50. https://doi.org/10.1002/wmon.1009 doi: 10.1002/wmon.1009
|
| [40] |
O. J. Schmitz, Fearful effects on ecological competitors, Nature, 570 (2019), 43–44. https://doi.org/10.1038/d41586-019-01712-7 doi: 10.1038/d41586-019-01712-7
|
| [41] |
B. L. Peckarsky, P. A. Abrams, D. I. Bolnick, L. M. Dill, J. H. Grabowski, B. Luttbeg, et al., Revisiting the classics: considering nonconsumptive effects in textbook examples of predator–prey interactions, Ecology, 89 (2008), 2416–2425. https://doi.org/10.1890/07-1131.1 doi: 10.1890/07-1131.1
|
| [42] | O. Schmitz, Predator and prey functional traits: understanding the adaptive machinery driving predator–prey interactions, F1000Res., 6 (2017). |
| [43] | J. Michaud, P. R. Barbosa, C. L. Bain, J. B. Torres, Extending the "ecology of fear" beyond prey: reciprocal nonconsumptive effects among competing aphid predators, Environ. Entomol., 45 (2016), 1398–1403. |
| [44] | M. H. Bayoumy, H. S. Awadalla, D. M. Fathy, T. M. Majerus, Beyond killing: intra-and interspecific nonconsumptive effects among aphidophagous competitors, Ecol. Entomol., 43 (2018), 794–803. |
| [45] |
P. Chesson, J. J. Kuang, The interaction between predation and competition, Nature, 456 (2008), 235–238. https://doi.org/10.1038/nature07248 doi: 10.1038/nature07248
|
| [46] | M. Arim, P. A. Marquet, Intraguild predation: a widespread interaction related to species biology, Ecol. Lett., 7 (2004), 557–564. |
| [47] | S.-E. Byun, S. Han, H. Kim, C. Centrallo, Us small retail businesses' perception of competition: Looking through a lens of fear, confidence, or cooperation, J. Retailing Consumer Serv., 52 (2020), 101925. |
| [48] |
Ø. Moen, The relationship between firm size, competitive advantages and export performance revisited, Int. Small Bus. J., 18 (1999), 53–72. https://doi.org/10.1177/0266242699181003 doi: 10.1177/0266242699181003
|
| [49] |
H. Löfgren, The communist party of India (marxist) and the left government in west Bengal, 1977–2011: Strains of governance and socialist imagination, Stud. Indian Polit., 4 (2016), 102–115. https://doi.org/10.1177/2321023016634947 doi: 10.1177/2321023016634947
|
| [50] | T. Wainwright, Narconomics: How to Run a Drug Cartel, PublicAffairs, 2016. |
| [51] | J. P. Sullivan, R. J. Bunker, Drug cartels, street gangs, and warlords, Small Wars Insurgencies, 13 (2002), 40–53. https://doi.org/10.1080/09592310208559180 |
| [52] | J. D. Murray, Mathematical Biology, Springer, 1993. https://doi.org/10.1007/978-3-662-08542-4 |
| [53] | R. S. Cantrell, C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, John Wiley & Sons, 2004. |
| [54] | A. Okubo, S. A. Levin, Diffusion and Ecological Problems: Modern Perspectives, Springer, 2001. https://doi.org/10.1007/978-1-4757-4978-6 |
| [55] |
J. Dockery, V. Hutson, K. Mischaikow, M. Pernarowski, The evolution of slow dispersal rates: a reaction diffusion model, J. Math. Biol., 37 (1998), 61–83. https://doi.org/10.1007/s002850050120 doi: 10.1007/s002850050120
|
| [56] |
X. He, W. M. Ni, The effects of diffusion and spatial variation in Lotka–Volterra competition–diffusion system I: heterogeneity vs. homogeneity, J. Differ. Equations, 254 (2013), 528–546. https://doi.org/10.1016/j.jde.2012.08.032 doi: 10.1016/j.jde.2012.08.032
|
| [57] |
A. Hastings, Can spatial variation alone lead to selection for dispersal?, Theor. Popul. Biol., 24 (1983), 244–251. https://doi.org/10.1016/0040-5809(83)90027-8 doi: 10.1016/0040-5809(83)90027-8
|
| [58] |
X. He, W. M. Ni, The effects of diffusion and spatial variation in Lotka–Volterra competition–diffusion system II: the general case, J. Differ. Equations, 254 (2013), 4088–4108. https://doi.org/10.1016/j.jde.2013.02.009 doi: 10.1016/j.jde.2013.02.009
|
| [59] |
X. He, W. M. Ni, Global dynamics of the Lotka–Volterra competition–diffusion system with equal amount of total resources, II, Cal. Var. Partial Differ. Equations, 55 (2016), 1–20. https://doi.org/10.1007/s00526-015-0942-y doi: 10.1007/s00526-015-0942-y
|
| [60] | H. Ninomiya, Separatrices of competition-diffusion equations, in Reaction-diffusion Equations And Their Applications And Computational Aspects-Proceedings Of The China-Japan Symposium, 1997,118. |
| [61] |
K. Y. Lam, W. M. Ni, Uniqueness and complete dynamics in heterogeneous competition-diffusion systems, SIAM J. Appl. Math., 72 (2012), 1695–1712. https://doi.org/10.1137/120869481 doi: 10.1137/120869481
|
| [62] |
Y. Lou, On the effects of migration and spatial heterogeneity on single and multiple species, J. Differ. Equations, 223 (2006), 400–426. https://doi.org/10.1016/j.jde.2005.05.010 doi: 10.1016/j.jde.2005.05.010
|
| [63] |
Y. Lou, S. Martínez, P. Poláčik, Loops and branches of coexistence states in a Lotka–Volterra ompetition model, J. Differ. Equations, 230 (2006), 720–742. https://doi.org/10.1016/j.jde.2006.04.005 doi: 10.1016/j.jde.2006.04.005
|
| [64] | W. M. Ni, Complete dynamics in a heterogeneous competition-diffusion system, East China Normal Univ. Univ. Minnesota, 2012 (2012). |
| [65] | L. Perko, Differential Equations and Dynamical Systems, Springer Science & Business Media, 2013. |
| [66] | D. Henry, Geometric Theory of Semilinear Parabolic Equations, Springer, 2006. |
| [67] | J. Morgan, Global existence for semilinear parabolic systems, SIAM J. Math. Anal., 20 (1989), 1128–1144. |
| [68] |
M. Pierre, Global existence in reaction-diffusion systems with control of mass: a survey, Milan J. Math., 78 (2010), 417–455. https://doi.org/10.1007/s00032-010-0133-4 doi: 10.1007/s00032-010-0133-4
|
| [69] |
K. Kishimoto, H. F. Weinberger, The spatial homogeneity of stable equilibria of some reaction-diffusion systems on convex domains, J. Differ. Equations, 58 (1985), 15–21. https://doi.org/10.1016/0022-0396(85)90020-8 doi: 10.1016/0022-0396(85)90020-8
|
| [70] | S. K. Sasmal, Y. Takeuchi, Dynamics of a predator-prey system with fear and group defense, J. Math. Anal. Appl., 481 (2020), 123471. |
| [71] |
Y. Du, Effects of a degeneracy in the competition model: part I. classical and generalized steady-state solutions, J. Differ. Equations, 181 (2002), 92–132. https://doi.org/10.1006/jdeq.2001.4074 doi: 10.1006/jdeq.2001.4074
|
| [72] |
Y. Du, Effects of a degeneracy in the competition model: Part II. perturbation and dynamical behaviour, J. Differ. Equations, 181 (2002), 133–164. https://doi.org/10.1006/jdeq.2001.4075 doi: 10.1006/jdeq.2001.4075
|
| [73] | D. Gilbarg, N. S. Trudinger, Elliptic Partial Differential Equations of Second Order, Springer, 1977. |
| [74] |
M. Krupa, M. Wechselberger, Local analysis near a folded saddle-node singularity, J. Differ. Equations, 248 (2010), 2841–2888. https://doi.org/10.1016/j.jde.2010.02.006 doi: 10.1016/j.jde.2010.02.006
|
| [75] |
V. Hajnová, L. Přibylová, Bifurcation manifolds in predator–prey models computed by Gröbner basis method, Math. Biosci., 312 (2019), 1–7. https://doi.org/10.1016/j.mbs.2019.03.008 doi: 10.1016/j.mbs.2019.03.008
|
| [76] |
L. van Veen, M. Hoti, Automatic detection of saddle-node–transcritical interactions, Int. J. Bifurcation Chaos, 29 (2019), 1950104. https://doi.org/10.1142/S0218127419501049 doi: 10.1142/S0218127419501049
|
| [77] |
A. Ekner, P. Tryjanowski, Do small hole nesting passerines detect cues left by a predator? A test on winter roosting sites, Acta Ornithol., 43 (2008), 107–111. https://doi.org/10.3161/000164508X345392 doi: 10.3161/000164508X345392
|
| [78] | N. Rodríguez, Q. Wang, L. Zhang, Understanding the effects of on-and off-hotspot policing: Evidence of hotspot, oscillating, and chaotic activities, SIAM J. Appl. Dyn. Syst., 20 (2021), 1882–1916. |
| [79] |
H. Berestycki, N. Rodriguez, L. Ryzhik, Traveling wave solutions in a reaction-diffusion model for criminal activity, Multiscale Model. Simul., 11 (2013), 1097–1126. https://doi.org/10.1137/12089884X doi: 10.1137/12089884X
|
| [80] |
R. D. Parshad, K. Antwi-Fordjour, E. M. Takyi, Some novel results in two species competition, SIAM J. Appl. Math., 81 (2021), 1847–1869. https://doi.org/10.1137/20M1387274 doi: 10.1137/20M1387274
|
| [81] |
R. D. Parshad, S. Wickramsooriya, S. Bailey, A remark on "biological control through provision of additional food to predators: A theoretical study", Theor. Popul. Biol., 132 (2020), 60–68. https://doi.org/10.1016/j.tpb.2007.03.011 doi: 10.1016/j.tpb.2007.03.011
|
| [82] |
A. P. Farrell, J. P. Collins, A. L. Greer, H. R. Thieme, Do fatal infectious diseases eradicate host species?, J. Math. Biol., 77 (2018), 2103–2164. https://doi.org/10.1007/s00285-018-1249-3 doi: 10.1007/s00285-018-1249-3
|
| [83] |
D. L. DeAngelis, W. M. Ni, B. Zhang, Dispersal and spatial heterogeneity: single species, J. Math. Biol., 72 (2016), 239–254. https://doi.org/10.1007/s00285-015-0879-y doi: 10.1007/s00285-015-0879-y
|
| [84] |
G. Wang, X. G. Liang, F. Z. Wang, The competitive dynamics of populations subject to an Allee effect, Ecol. Model., 124 (1999), 183–192. https://doi.org/10.1016/S0304-3800(99)00160-X doi: 10.1016/S0304-3800(99)00160-X
|
| [85] |
M. De Silva, S. R. J. Jang, Competitive exclusion and coexistence in a Lotka–Volterra competition model with Allee effects and stocking, Lett. Biomath., 2 (2015), 56–66. https://doi.org/10.30707/LiB2.1DeSilva doi: 10.30707/LiB2.1DeSilva
|
| [86] |
P. A. Naik, Z. Eskandari, M. Yavuz, J. Zu, Complex dynamics of a discrete-time Bazykin–Berezovskaya prey-predator model with a strong Allee effect, J. Comput. Appl. Math., 413 (2022), 114401. https://doi.org/10.1016/j.cam.2022.114401 doi: 10.1016/j.cam.2022.114401
|
| [87] |
M. Yavuz, N. Sene, Stability analysis and numerical computation of the fractional predator–prey model with the harvesting rate, Fractal Fract., 4 (2020), 35. https://doi.org/10.3390/fractalfract4030035 doi: 10.3390/fractalfract4030035
|
| [88] |
Z. Hammouch, M. Yavuz, N. Özdemir, Numerical solutions and synchronization of a variable-order fractional chaotic system, Math. Model. Numer. Simul. Appl., 1 (2021), 11–23. https://doi.org/10.53391/mmnsa.2021.01.002 doi: 10.53391/mmnsa.2021.01.002
|
| [89] |
M. Gholami, R. K. Ghaziani, Z. Eskandari, Three-dimensional fractional system with the stability condition and chaos control, Math. Model. Numer. Simul. Appl., 2 (2022), 41–47. https://doi.org/10.53391/mmnsa.2022.01.004 doi: 10.53391/mmnsa.2022.01.004
|
| [90] | H. Joshi, B. K. Jha, Chaos of calcium diffusion in Parkinson's infectious disease model and treatment mechanism via Hilfer fractional derivative, Math. Model. Numer. Simul. Appl., 1 (2021), 84–94. |
| [91] |
I. Mazari, G. Nadin, Y. Privat, Optimal location of resources maximizing the total population size in logistic models, J. Math. Pures Appl., 134 (2020), 1–35. https://doi.org/10.1016/j.matpur.2019.10.008 doi: 10.1016/j.matpur.2019.10.008
|
| [92] |
D. DeAngelis, B. Zhang, W. M. Ni, Y. Wang, Carrying capacity of a population diffusing in a heterogeneous environment, Mathematics, 8 (2020), 49. https://doi.org/10.3390/math8010049 doi: 10.3390/math8010049
|
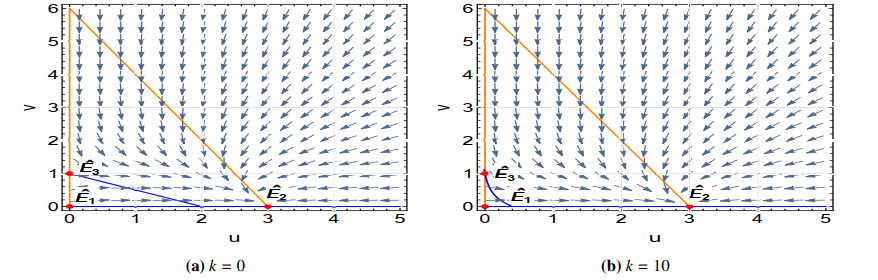
















Figures(17) / Tables(1)

Vaibhava Srivastava, Eric M. Takyi, Rana D. Parshad. The effect of "fear" on two species competition[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 8814-8855. doi: 10.3934/mbe.2023388







 DownLoad:
DownLoad: