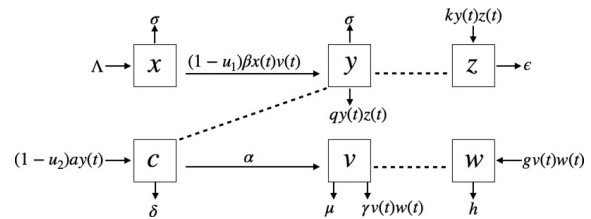
In this paper, a mathematical model describing the hepatitis B virus (HBV) infection of hepatocytes with the intracellular HBV-DNA containing capsids, cytotoxic T-lymphocyte (CTL), antibodies including drug therapy (blocking new infection and inhibiting viral production) with two-time delays is studied. It incorporates the delay in the productively infected hepatocytes and the delay in an antigenic stimulation generating CTL. We verify the positivity and boundedness of solutions and determine the basic reproduction number. The local and global stability of three equilibrium points (infection-free, immune-free, and immune-activated) are investigated. Finally, the numerical simulations are established to show the role of these therapies in reducing viral replication and HBV infection. Our results show that the treatment by blocking new infection gives more significant results than the treatment by inhibiting viral production for infected hepatocytes. Further, both delays affect the number of infections and duration i.e. the longer the delay, the more severe the HBV infection.
Citation: Pensiri Yosyingyong, Ratchada Viriyapong. Global dynamics of multiple delays within-host model for a hepatitis B virus infection of hepatocytes with immune response and drug therapy[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 7349-7386. doi: 10.3934/mbe.2023319
In this paper, a mathematical model describing the hepatitis B virus (HBV) infection of hepatocytes with the intracellular HBV-DNA containing capsids, cytotoxic T-lymphocyte (CTL), antibodies including drug therapy (blocking new infection and inhibiting viral production) with two-time delays is studied. It incorporates the delay in the productively infected hepatocytes and the delay in an antigenic stimulation generating CTL. We verify the positivity and boundedness of solutions and determine the basic reproduction number. The local and global stability of three equilibrium points (infection-free, immune-free, and immune-activated) are investigated. Finally, the numerical simulations are established to show the role of these therapies in reducing viral replication and HBV infection. Our results show that the treatment by blocking new infection gives more significant results than the treatment by inhibiting viral production for infected hepatocytes. Further, both delays affect the number of infections and duration i.e. the longer the delay, the more severe the HBV infection.

| [1] |
R. M. Ribeiro, A. Lo, A. S. Perelson, Dynamics of hepatitis b virus infection, Microb. Infect., 4 (2002), 829–835, https://doi.org/10.1016/S1286-4579(02)01603-9 doi: 10.1016/S1286-4579(02)01603-9

|
| [2] | World Health Organization, Hepatitis B, 2020, Available from: http://www.who.int/mediacentre/factsheets/fs204/en |
| [3] |
L. V. Tsui, L. G. Guidotti, T. Ishikawa, F. V. Chisari, Posttranscriptional clearance of hepatitis B virus RNA by cytotoxic t lymphocyte-activated hepatocytes, Proceed. Nat. Acad. Sci., 92 (1995), 12398–12402. https://doi.org/10.1073/pnas.92.26.12398 doi: 10.1073/pnas.92.26.12398
|
| [4] |
L. G. Guidotti, T. Ishikawa, M. V. Hobbs, B. Matzke, R. Schreiber, F. V. Chisari, Intracellular inactivation of the hepatitis B virus by cytotoxic t lymphocytes, Immunity, 4 (1996), 25–36. https://doi.org/10.1016/S1074-7613(00)80295-2 doi: 10.1016/S1074-7613(00)80295-2
|
| [5] |
L. G. Guidotti, F. V. Chisari, To kill or to cure: Options in host defense against viral infection, Current Opin. Immunol., 8 (1996), 478–483. https://doi.org/10.1016/S0952-7915(96)80034-3 doi: 10.1016/S0952-7915(96)80034-3
|
| [6] |
L. G. Guidotti, R. Rochford, J. Chung, M. Shapiro, R. Purcell, F. V. Chisari, Viral clearance without destruction of infected cells during acute hbv infection, Science, 284 (1999), 825–829. https://doi.org/10.1126/science.284.5415.825 doi: 10.1126/science.284.5415.825
|
| [7] |
M. Iannacone, G. Sitia, L. G. Guidotti, Pathogenetic and antiviral immune responses against hepatitis b virus, Future Virol., 1 (2006), 189–196. https://doi.org/10.2217/17460794.1.2.189 doi: 10.2217/17460794.1.2.189
|
| [8] | C. Long, H. Qi, S.-H. Huang, Mathematical modeling of cytotoxic lymphocyte-mediated immune response to hepatitis B virus infection, J. Biomed. Biotechnol., 2008. https: //doi.org/10.1155/2008/743690 |
| [9] |
S. Phillips, S. Chokshi, A. Riva, A. Evans, R. Williams, N. V. Naoumov, Cd8+ T cell control of hepatitis B virus replication: Direct comparison between cytolytic and noncytolytic functions, J. Immunol., 184 (2010), 287–295. https://doi.org/10.4049/jimmunol.0902761 doi: 10.4049/jimmunol.0902761
|
| [10] |
F. V. Chisari, M. Isogawa, S. F. Wieland, Pathogenesis of hepatitis b virus infection, Pathol. Biol., 58 (2010), 258–266. https://doi.org/10.1016/j.patbio.2009.11.001 doi: 10.1016/j.patbio.2009.11.001
|
| [11] |
A. Busca, A. Kumar, Innate immune responses in hepatitis B virus (HBV) infection, Virol. J., 11 (2014), 1–8, https://doi.org/10.1186/1743-422X-11-22 doi: 10.1186/1743-422X-11-22
|
| [12] |
A. Elaiw, N. Alshamrani, Global analysis for a delay-distributed viral infection model with antibodies and general nonlinear incidence rate, J. Korean Soc. Indust. Appl. Math., 18 (2014), 317–335. https://doi.org/10.12941/jksiam.2014.18.317 doi: 10.12941/jksiam.2014.18.317
|
| [13] |
Y. Wang, Y. Zhou, J. Wu, J. Heffernan, Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104–112. https://doi.org/10.1016/j.mbs.2009.03.003 doi: 10.1016/j.mbs.2009.03.003
|
| [14] |
S. Wang, X. Song, Z. Ge, Dynamics analysis of a delayed viral infection model with immune impairment, Appl. Math. Model., 35 (2011), 4877–4885. https://doi.org/10.1016/j.apm.2011.03.043 doi: 10.1016/j.apm.2011.03.043
|
| [15] |
N. Bairagi, D. Adak, Global analysis of HIV-1 dynamics with hill type infection rate and intracellular delay, Appl. Math. Model., 38 (2014), 5047–5066. https://doi.org/10.1016/j.apm.2014.03.010 doi: 10.1016/j.apm.2014.03.010
|
| [16] |
B.-Z. Guo, L.-M. Cai, A note for the global stability of a delay differential equation of hepatitis B virus infection, Math. Biosci. Eng., 8 (2011), 689. https://doi.org/10.3934/mbe.2011.8.689 doi: 10.3934/mbe.2011.8.689
|
| [17] |
H. Song, W. Jiang, S. Liu, Virus dynamics model with intracellular delays and immune response, Math. Biosci. Eng., 12 (2015), 185. https://doi.org/10.3934/mbe.2015.12.185 doi: 10.3934/mbe.2015.12.185
|
| [18] |
A. Meskaf, K. Allali, Y. Tabit, Optimal control of a delayed hepatitis B viral infection model with cytotoxic T-lymphocyte and antibody responses, Int. J. Dynam. Control, 5 (2017), 893–902. https://doi.org/10.1007/s40435-016-0231-4 doi: 10.1007/s40435-016-0231-4
|
| [19] |
K. Manna, S. P. Chakrabarty, Global stability of one and two discrete delay models for chronic hepatitis B infection with HBV DNA-containing capsids, Comput. Appl. Math., 36 (2017), 525–536. https://doi.org/10.1007/s40314-015-0242-3 doi: 10.1007/s40314-015-0242-3
|
| [20] |
J. Danane, K. Allali, Mathematical analysis and treatment for a delayed hepatitis B viral infection model with the adaptive immune response and DNA-containing capsids, High-throughput, 7 (2018), 35. https://doi.org/10.3390/ht7040035 doi: 10.3390/ht7040035
|
| [21] |
A. Bertoletti, M. K. Maini, C. Ferrari, The host-pathogen interaction during HBV infection: Immunological controversies, Antiviral Therapy, 15 (2010), 15–24. https://doi.org/10.3851/IMP1620 doi: 10.3851/IMP1620
|
| [22] |
S. J. Hadziyannis, N. C. Tassopoulos, E. J. Heathcote, T.-T. Chang, G. Kitis, M. Rizzetto, et al., Adefovir dipivoxil for the treatment of hepatitis B e antigen–negative chronic hepatitis B, New England J. Med., 348 (2003), 800–807. https://doi.org/10.1056/NEJMoa021812 doi: 10.1056/NEJMoa021812
|
| [23] |
L. L. Stein, R. Loomba, Drug targets in hepatitis b virus infection, Infect. Disorders-Drug Targets (Formerly Current Drug Targets-Infectious Disorders), 9 (2009), 105–116. https://doi.org/10.2174/187152609787847677 doi: 10.2174/187152609787847677
|
| [24] |
S. Hagiwara, N. Nishida, M. Kudo, Antiviral therapy for chronic hepatitis B: Combination of nucleoside analogs and interferon, World J. Hepatol., 7 (2015), 2427. https://doi.org/10.4254/wjh.v7.i23.2427 doi: 10.4254/wjh.v7.i23.2427
|
| [25] |
F. van den Berg, S. W. Limani, N. Mnyandu, M. B. Maepa, A. Ely, P. Arbuthnot, Advances with rnai-based therapy for hepatitis b virus infection, Viruses, 12 (2020), 851. https://doi.org/10.3390/v12080851 doi: 10.3390/v12080851
|
| [26] | J. S. Nayagam, Z. C. Cargill, K. Agarwal, The role of rna interference in functional cure strategies for chronic hepatitis B, Current Hepatol. Rep., 1–8. https: //doi.org/10.1007/s11901-020-00548-4 |
| [27] |
E. De Clercq, G. Férir, S. Kaptein, J. Neyts, Antiviral treatment of chronic hepatitis B virus (HBV) infections, Viruses, 2 (2010), 1279–1305. https://doi.org/10.3390/v2061279 doi: 10.3390/v2061279
|
| [28] |
G. K. Lau, T. Piratvisuth, K. X. Luo, P. Marcellin, S. Thongsawat, G. Cooksley, et al., Peginterferon alfa-2a, lamivudine, and the combination for hbeag-positive chronic hepatitis B, New England J. Med., 352 (2005), 2682–2695. https://doi.org/10.1056/NEJMoa043470 doi: 10.1056/NEJMoa043470
|
| [29] |
M. A. Nowak, S. Bonhoeffer, A. M. Hill, R. Boehme, H. C. Thomas, H. McDade, Viral dynamics in hepatitis b virus infection, Proceed. Nat. Acad. Sci., 93 (1996), 4398–4402. https://doi.org/10.1073/pnas.93.9.4398 doi: 10.1073/pnas.93.9.4398
|
| [30] |
A. Goyal, R. M. Ribeiro, A. S. Perelson, The role of infected cell proliferation in the clearance of acute HBV infection in humans, Viruses, 9 (2017), 350. https://doi.org/10.3390/v9110350 doi: 10.3390/v9110350
|
| [31] |
S. M. Ciupe, R. M. Ribeiro, P. W. Nelson, G. Dusheiko, A. S. Perelson, The role of cells refractory to productive infection in acute hepatitis b viral dynamics, Proceed. Nat. Acad. Sci., 104 (2007), 5050–5055. https://doi.org/10.1073/pnas.0603626104 doi: 10.1073/pnas.0603626104
|
| [32] |
S. M. Ciupe, R. M. Ribeiro, P. W. Nelson, A. S. Perelson, Modeling the mechanisms of acute hepatitis b virus infection, J. Theor. Biol., 247 (2007), 23–35. https://doi.org/10.1016/j.jtbi.2007.02.017 doi: 10.1016/j.jtbi.2007.02.017
|
| [33] |
S. Hews, S. Eikenberry, J. D. Nagy, Y. Kuang, Rich dynamics of a hepatitis b viral infection model with logistic hepatocyte growth, J. Math. Biol., 60 (2010), 573–590. https://doi.org/10.1007/s00285-009-0278-3 doi: 10.1007/s00285-009-0278-3
|
| [34] |
N. Yousfi, K. Hattaf, A. Tridane, Modeling the adaptive immune response in HBV infection, J. Math. Biol., 63 (2011), 933–957. https://doi.org/10.1007/s00285-010-0397-x doi: 10.1007/s00285-010-0397-x
|
| [35] |
A. M. Elaiw, N. Almuallem, Global properties of delayed-HIV dynamics models with differential drug efficacy in cocirculating target cells, Appl. Math. Comput., 265 (2015), 1067–1089. https://doi.org/10.1016/j.amc.2015.06.011 doi: 10.1016/j.amc.2015.06.011
|
| [36] |
K. Mboya, D. Makinde, E. Massawe, Cytotoxic cells and control strategies are effective in reducing the HBV infection through a mathematical modelling, Int. J. Prevent. Treatment, 4 (2015), 48–57. https://doi.org/10.5923/j.ijpt.20150403.02 doi: 10.5923/j.ijpt.20150403.02
|
| [37] | A. Tridane, K. Hattaf, R. Yafia, F. A. Rihan, Mathematical modeling of HBV with the antiviral therapy for the immunocompromised patients, Commun. Math. Biol. Neurosci., 2016 (2016). |
| [38] |
P. Yosyingyong, R. Viriyapong, Global stability and optimal control for a hepatitis b virus infection model with immune response and drug therapy, J. Appl. Math. Comput., 60 (2019), 537–565. https://doi.org/10.1007/s12190-018-01226-x doi: 10.1007/s12190-018-01226-x
|
| [39] |
K. Manna, K. Hattaf, A generalized distributed delay model for hepatitis b virus infection with two modes of transmission and adaptive immunity: A mathematical study, Math. Methods Appl. Sci., 45 (2022), 11614–11634. https://doi.org/10.1002/mma.8470 doi: 10.1002/mma.8470
|
| [40] |
S. R. Lewin, R. M. Ribeiro, T. Walters, G. K. Lau, S. Bowden, S. Locarnini, et al., Analysis of hepatitis b viral load decline under potent therapy: Complex decay profiles observed, Hepatology, 34 (2001), 1012–1020. https://doi.org/10.1053/jhep.2001.28509 doi: 10.1053/jhep.2001.28509
|
| [41] |
S. Eikenberry, S. Hews, J. D. Nagy, Y. Kuang, The dynamics of a delay model of hepatitis b virus infection with logistic hepatocyte growth, Math. Biosci. Eng., 6 (2009), 283. https://doi.org/10.3934/mbe.2009.6.283 doi: 10.3934/mbe.2009.6.283
|
| [42] |
S. A. Gourley, Y. Kuang, J. D. Nagy, Dynamics of a delay differential equation model of hepatitis b virus infection, J. Biol. Dynam., 2 (2008), 140–153. https://doi.org/10.1080/17513750701769873 doi: 10.1080/17513750701769873
|
| [43] |
Q. Xie, D. Huang, S. Zhang, J. Cao, Analysis of a viral infection model with delayed immune response, Appl. Math. Model., 34 (2010), 2388–2395. https://doi.org/10.1016/j.apm.2009.11.005 doi: 10.1016/j.apm.2009.11.005
|
| [44] |
K. Wang, W. Wang, S. Song, Dynamics of an HBV model with diffusion and delay, J. Theor. Biol., 253 (2008), 36–44, https://doi.org/10.1016/j.jtbi.2007.11.007 doi: 10.1016/j.jtbi.2007.11.007
|
| [45] |
P. Fisicaro, C. Valdatta, C. Boni, M. Massari, C. Mori, A. Zerbini, et al., Early kinetics of innate and adaptive immune responses during hepatitis b virus infection, Gut, 58 (2009), 974–982. http://dx.doi.org/10.1136/gut.2008.163600 doi: 10.1136/gut.2008.163600
|
| [46] | D. Sun, F. Liu, Analysis of a new delayed HBV model with exposed state and immune response to infected cells and viruses, BioMed Res. Int., 2017. https: //doi.org/10.1155/2017/7805675 |
| [47] |
K. Manna, S. P. Chakrabarty, Chronic hepatitis b infection and HBV DNA-containing capsids: Modeling and analysis, Commun. Nonlinear Sci. Numer. Simul., 22 (2015), 383–395. https://doi.org/10.1016/j.cnsns.2014.08.036 doi: 10.1016/j.cnsns.2014.08.036
|
| [48] |
T. Guo, H. Liu, C. Xu, F. Yan, Global stability of a diffusive and delayed HBV infection model with HBV DNA-containing capsids and general incidence rate, Discrete Continuous Dynam. Systems-B, 23 (2018), 4223. https://doi.org/10.3934/dcdsb.2018134 doi: 10.3934/dcdsb.2018134
|
| [49] |
M. Aniji, N. Kavitha, S. Balamuralitharan, Mathematical modeling of hepatitis b virus infection for antiviral therapy using lham, Adv. Differ. Equat., 2020 (2020), 1–19. https://doi.org/10.1186/s13662-020-02770-2 doi: 10.1186/s13662-020-02770-2
|
| [50] | K. Allali, A. Meskaf, A. Tridane, Mathematical modeling of the adaptive immune responses in the early stage of the HBV infection, Int. J. Differ. Equat., 2018. https: //doi.org/10.1155/2018/6710575 |
| [51] | N. Chan Chí, E. AvilaVales, G. García Almeida, Analysis of a HBV model with diffusion and time delay, J. Appl. Math., 2012. https: //doi.org/10.1155/2012/578561 |
| [52] |
K. Hattaf, N. Yousfi, A generalized HBV model with diffusion and two delays, Comput. Math. Appl., 69 (2015), 31–40. https://doi.org/10.1016/j.camwa.2014.11.010 doi: 10.1016/j.camwa.2014.11.010
|
| [53] | K. Hattaf, M. Rachik, S. Saadi, N. Yousfi, Optimal control of treatment in a basic virus infection model, Appl. Math. Sci., 3 (2009), 949–958. |
| [54] |
K. Manna, S. P. Chakrabarty, Combination therapy of pegylated interferon and lamivudine and optimal controls for chronic hepatitis b infection, Int. J. Dynam. Control, 6 (2018), 354–368. https://doi.org/10.1007/s40435-017-0306-x doi: 10.1007/s40435-017-0306-x
|
| [55] |
K. Hattaf, A new generalized definition of fractional derivative with non-singular kernel, Computation, 8 (2020), 49. https://doi.org/10.3390/computation8020049 doi: 10.3390/computation8020049
|
| [56] |
K. Hattaf, On the stability and numerical scheme of fractional differential equations with application to biology, Computation, 10 (2022), 97. https://doi.org/10.3390/computation10060097 doi: 10.3390/computation10060097
|
| [57] |
S. M. Ciupe, R. M. Ribeiro, A. S. Perelson, Antibody responses during hepatitis b viral infection, PLoS Comput. Biol., 10 (2014), e1003730. https://doi.org/10.1371/journal.pcbi.1003730 doi: 10.1371/journal.pcbi.1003730
|
| [58] |
R. Ahmed, D. Gray, Immunological memory and protective immunity: Understanding their relation, Science, 272 (1996), 54–60. https://doi.org/10.1126/science.272.5258 doi: 10.1126/science.272.5258
|
| [59] | J. K. Hale, S. M. V. Lunel, Introduction to functional differential equations, vol. 99, Springer Science & Business Media, 2013. |
| [60] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|







Figures(8) / Tables(1)

Pensiri Yosyingyong, Ratchada Viriyapong. Global dynamics of multiple delays within-host model for a hepatitis B virus infection of hepatocytes with immune response and drug therapy[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 7349-7386. doi: 10.3934/mbe.2023319







 DownLoad:
DownLoad: