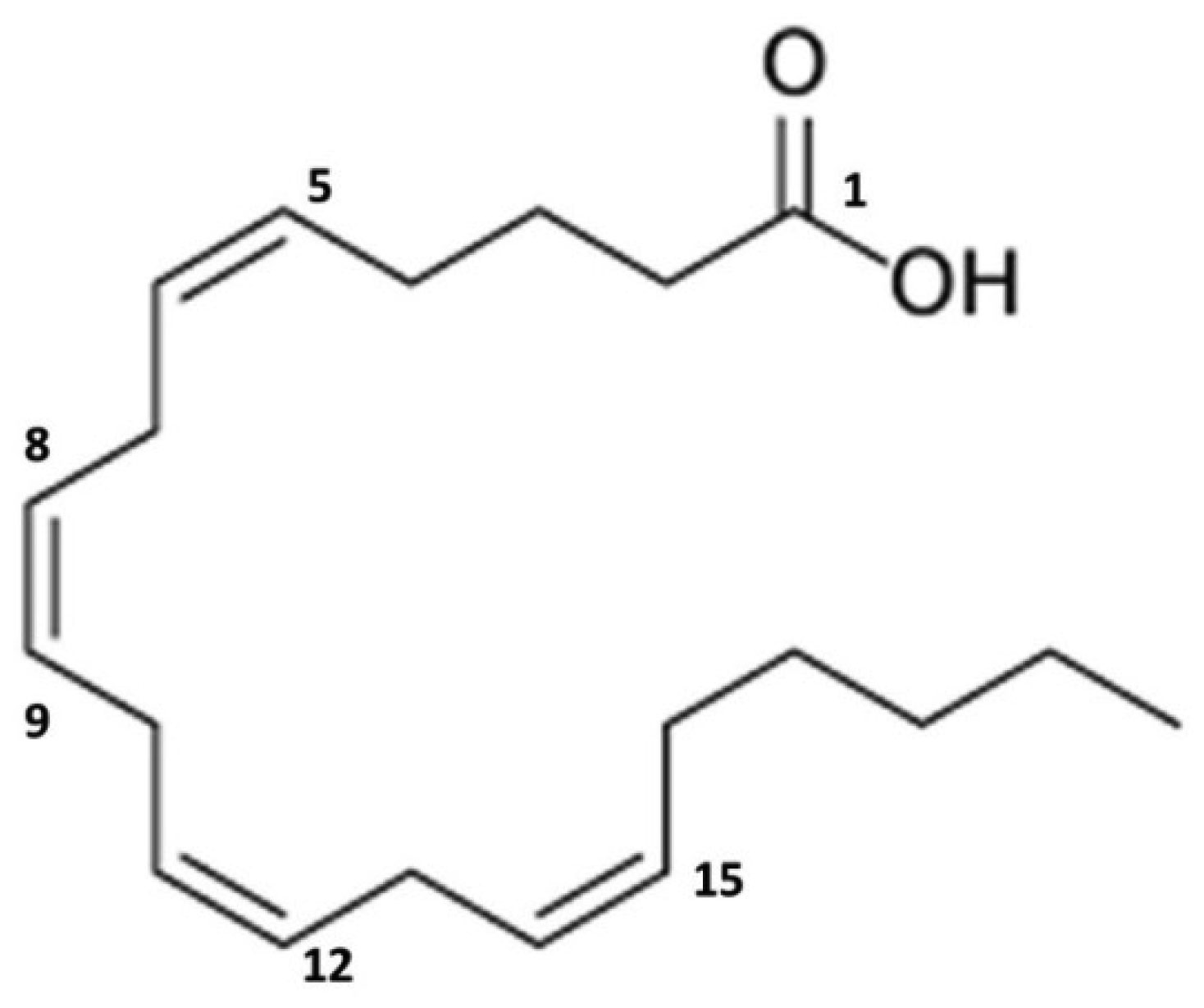
Mast cells and basophils degranulate upon activation, releasing preformed mediators from intracellular granules into the extracellular environment, of which tryptase and histamine are the two most common and best characterized mediators. Despite the large number of mediators synthesized by mast cells, the non-tryptase biomarkers used to evaluate systemic mastocytosis and mast cell activation syndrome do not include the metabolites of the prestored amine histamine and the de novo synthesized phospholipids prostaglandin D2 and leukotriene C4. Currently, these markers are not used as criteria for the diagnosis of mastocytosis and mast cell activation syndrome. However, consensus groups foster the use of increases in measured baseline levels of these metabolites as potential diagnostic criteria. Metabolites of arachidonic acid such as prostaglandin D2 or leukotriene C4 play a role in the development of symptoms in systemic mastocytosis and mast cell activation syndrome. In this review, the metabolites of arachidonic acid and the detection of the metabolites of leukotrienes and prostaglandins in mastocytosis are highlighted. Measurement of these metabolites remains a major challenge because they are not widely available in daily clinical practice. However, new insights have been gained in recent years, and their application in the clinic has progressed.
Citation: Peter Jandus. Arachidonic acid metabolism and its use in the diagnosis of mastocytosis[J]. AIMS Allergy and Immunology, 2023, 7(1): 82-91. doi: 10.3934/Allergy.2023006
Mast cells and basophils degranulate upon activation, releasing preformed mediators from intracellular granules into the extracellular environment, of which tryptase and histamine are the two most common and best characterized mediators. Despite the large number of mediators synthesized by mast cells, the non-tryptase biomarkers used to evaluate systemic mastocytosis and mast cell activation syndrome do not include the metabolites of the prestored amine histamine and the de novo synthesized phospholipids prostaglandin D2 and leukotriene C4. Currently, these markers are not used as criteria for the diagnosis of mastocytosis and mast cell activation syndrome. However, consensus groups foster the use of increases in measured baseline levels of these metabolites as potential diagnostic criteria. Metabolites of arachidonic acid such as prostaglandin D2 or leukotriene C4 play a role in the development of symptoms in systemic mastocytosis and mast cell activation syndrome. In this review, the metabolites of arachidonic acid and the detection of the metabolites of leukotrienes and prostaglandins in mastocytosis are highlighted. Measurement of these metabolites remains a major challenge because they are not widely available in daily clinical practice. However, new insights have been gained in recent years, and their application in the clinic has progressed.

| [1] |
Krystel-Whittemore M, Dileepan KN, Wood JG (2016) Mast cell: A multi-functional master cell. Front Immunol 6: 620. https://doi.org/10.3389/fimmu.2015.00620 
|
| [2] |
Theoharides TC, Tsilioni I, Ren H (2019) Recent advances in our understanding of mast cell activation-or should it be mast cell mediator disorders?. Expert Rev Clin Immunol 15: 639-656. https://doi.org/10.1080/1744666X.2019.1596800
|
| [3] |
Valent P, Hartmann K, Bonadonna P, et al. (2022) Global classification of mast cell activation disorders: An ICD-10-CM-adjusted proposal of the ECNM-AIM consortium. J Allergy Clin Immunol Pract 10: 1941-1950. https://doi.org/10.1016/j.jaip.2022.05.007
|
| [4] |
Gülen T, Akin C, Bonadonna P, et al. (2021) Selecting the right criteria and proper classification to diagnose mast cell activation syndromes: A critical review. J Allergy Clin Immunol Pract 9: 3918-3928. https://doi.org/10.1016/j.jaip.2021.06.011
|
| [5] |
Paivandy A, Pejler G (2021) Novel strategies to target mast cells in disease. J Innate Immun 13: 131-147. https://doi.org/10.1159/000513582
|
| [6] | Rabinovitch N (2007) Urinary leukotriene E4. Immunol Allergy Clin 27: 651-664. https://doi.org/10.1016/j.iac.2007.09.004 |
| [7] |
Roberts II JL, Sweetman BJ, Lewis RA, et al. (1980) Increased production of prostaglandin D2 in patients with systemic mastocytosis. N Engl J Med 303: 1400-1404. https://doi.org/10.1056/NEJM198012113032405
|
| [8] |
Valent P, Akin C, Arock M, et al. (2012) Definitions, criteria and global classification of mast cell disorders with special reference to mast cell activation syndromes: a consensus proposal. Int Arch Allergy Imm 157: 215-225. https://doi.org/10.1159/000328760
|
| [9] |
Valent P, Hartmann K, Schwaab J, et al. (2022) Personalized management strategies in mast cell disorders: ECNM-AIM user's guide for daily clinical practice. J Allergy Clin Immunol Pract 10: 1999-2012. https://doi.org/10.1016/j.jaip.2022.03.007
|
| [10] |
Hanna VS, Hafez EAA (2018) Synopsis of arachidonic acid metabolism: A review. J Adv Res 11: 23-32. https://doi.org/10.1016/j.jare.2018.03.005
|
| [11] |
Wang B, Wu L, Chen J, et al. (2021) Metabolism pathways of arachidonic acids: mechanisms and potential therapeutic targets. Signal Transduct Target Ther 6: 94. https://doi.org/10.1038/s41392-020-00443-w
|
| [12] |
Yokoyama C, Tanabe T (1989) Cloning of human gene encoding prostaglandin endoperoxide synthase and primary structure of the enzyme. Biochem Biophys Res Commun 165: 888-894. https://doi.org/10.1016/S0006-291X(89)80049-X
|
| [13] |
Funk CD, Funk LB, Kennedy ME, et al. (1991) Human platelet/erythroleukemia cell prostaglandin G/H synthase: cDNA cloning, expression, and gene chromosomal assignment. FASEB J 5: 2304-2312. https://doi.org/10.1096/fasebj.5.9.1907252
|
| [14] |
Hla T, Neilson K (1992) Human cyclooxygenase-2 cDNA. P Natl Acad Sci USA 89: 7384-7388. https://doi.org/10.1073/pnas.89.16.7384
|
| [15] |
Mulugeta S, Suzuki T, Hernandez NT, et al. (2010) Identification and absolute configuration of dihydroxy-arachidonic acids formed by oxygenation of 5S-HETE by native and aspirin-acetylated COX-2. J Lipid Res 51: 575-585. https://doi.org/10.1194/jlr.M001719
|
| [16] |
Chavis C, Vachier I, Godard P, et al. (2000) Lipoxins and other arachidonate derived mediators in bronchial asthma. Thorax 55: S38-S41. https://doi.org/10.1136/thorax.55.suppl_2.S38
|
| [17] |
Serhan CN (2005) Lipoxins and aspirin-triggered 15-epi-lipoxins are the first lipid mediators of endogenous anti-inflammation and resolution. Prostag Leukotr Ess 73: 141-162. https://doi.org/10.1016/j.plefa.2005.05.002
|
| [18] |
Ugajin T, Satoh T, Kanamori T, et al. (2011) FcϵRI, but not FcγR, signals induce prostaglandin D2 and E2 production from basophils. Am J Pathol 179: 775-782. https://doi.org/10.1016/j.ajpath.2011.04.023
|
| [19] |
Urade Y, Hayashi O (2000) Biochemical, structural, genetic, physiological, and pathophysiological features of lipocalin-type prostaglandin D synthase. Biochim Biophys Acta 1482: 259-271. https://doi.org/10.1016/S0167-4838(00)00161-8
|
| [20] |
Song WL, Wang M, Riccioitti E, et al. (2008) Tetranor PGDM, an abundant urinary metabolite reflects synthesis of PGD2 in mice and humans. J Biol Chem 283: 1179-1188. https://doi.org/10.1074/jbc.M706839200
|
| [21] |
Liston TE, Roberts II LJ (1985) Transformation of prostaglandin D2 to 9 alpha, 11 beta-(15S)-trihydroxyprosta-(5Z,13E)-dien-1-oic acid (9 alpha, 11 beta-prostaglandin F2): a unique biologically active prostaglandin produced enzymatically in vivo in humans. P Natl Acad Sci USA 82: 6030-6034. https://doi.org/10.1073/pnas.82.18.6030
|
| [22] |
Cho C, Nguyen A, Bryant KJ, et al. (2016) Prostaglandin D2 metabolites as a biomarker of in vivo mast cell activation in systemic mastocytosis and rheumatoid arthritis. Immun Inflamm Dis 4: 64-69. https://doi.org/10.1002/iid3.94
|
| [23] |
Roberts II LJ, Oates JA (1991) Biochemical diagnosis of systemic mast cell disorders. J Invest Dermatol 96: 19S-25S. https://doi.org/10.1111/1523-1747.ep12468945
|
| [24] | Kootte AMM, Haak A, Roberts II LJ (1983) The flush syndrome: an expression of systemic mastocytosis with increased prostaglandin D2 production. Neth J Med 26: 18-20. |
| [25] | Roberts II LJ, Fields JP, Oates JA (1982) Mastocytosis without urticaria pigmentosa: A frequently unrecognized cause of recurrent syncope. Trans Assoc Am Physicians 95: 36-41. |
| [26] |
Denzlinger C, Haberl C, Wilmanns W (1995) Cysteinyl leukotriene production in anaphylactic reactions. Int Arch Allergy Imm 108: 158-164. https://doi.org/10.1159/000237133
|
| [27] |
Ono E, Taniguchi M, Mita H, et al. (2009) Increased production of cysteinyl leukotrienes and prostaglandin D2 during human anaphylaxis. Clin Exp Allergy 39: 72-80. https://doi.org/10.1111/j.1365-2222.2008.03104.x
|
| [28] |
Mita H, Endoh S, Kudoh M, et al. (2001) Possible involvement of mast-cell activation in aspirin provocation of aspirin-induced asthma. Allergy 56: 1061-1067. https://doi.org/10.1111/j.1398-9995.2001.00913.x
|
| [29] |
Brannan JD, Gulliksson M, Anderson SD, et al. (2003) Evidence of mast cell activation and leukotriene release after mannitol inhalation. Eur Respir J 22: 491-496. https://doi.org/10.1183/09031936.03.00113403
|
| [30] |
Awad JA, Morrow JD, Roberts II LJ (1994) Detection of the major urinary metabolite of prostaglandin D2 in the circulation: Demonstration of elevated levels in patients with disorders of systemic mast cell activation. J Allergy Clin Immunol 93: 817-824. https://doi.org/10.1016/0091-6749(94)90371-9
|
| [31] |
Ravi A, Butterfield JH, Weiler CR (2014) Mast cell activation syndrome: improved identification by combined determinations of serum tryptase and 24 hour urine 11beta prostaglandin2alpha. J Allergy Clin Immunol Pract 2: 775-778. https://doi.org/10.1016/j.jaip.2014.06.011
|
| [32] |
Morrow JD, Guzzo C, Lazarus G, et al. (1995) Improved diagnosis of mastocytosis by measurement of the major urinary metabolite of prostaglandin D2. J Invest Dermatol 104: 937-940. https://doi.org/10.1111/1523-1747.ep12606209
|
| [33] |
Butterfield JH (2020) Survey of mast cell mediator levels from patients presenting with symptoms of mast cell activation. Int Arch Allergy Imm 181: 43-50. https://doi.org/10.1159/000503964
|
| [34] |
Divekar R, Butterfield J (2015) Urinary 11β-PGF2α and N-methyl histamine correlate with bone marrow biopsy findings in mast cell disorders. Allergy 70: 1230-1238. https://doi.org/10.1111/all.12668
|
| [35] |
Butterfield JH, Weiler CR (2008) Prevention of mast cell activation disorder-associated clinical sequelae of excessive prostaglandin D2 production. Int Arch Allergy Imm 147: 338-343. https://doi.org/10.1159/000144042
|
| [36] |
Friedman BS, Steinberg SC, Meggs WJ, et al. (1989) Analysis of plasma histamine levels in patients with mast cell disorders. Am J Med 87: 649-654. https://doi.org/10.1016/S0002-9343(89)80398-5
|
| [37] |
Kanaoka Y, Austen KF (2019) Roles of cysteinyl leukotrienes and their receptors in immune cell-related functions. Adv Immunol 142: 65-84. https://doi.org/10.1016/bs.ai.2019.04.002
|
| [38] |
MacGlashan DW, Schleimer RP, Peters SP, et al. (1982) Generation of leukotrienes by purified human lung mast cells. J Clin Invest 70: 747-751. https://doi.org/10.1172/JCI110670
|
| [39] | MacGlashan DW, Schleimer RP, Peters SP, et al. (1983) Comparative studies of human basophils and mast cells. Fed Proc 42: 2504-2509. |
| [40] | Schulman ES, MacGlashan DW, Schleimer RP, et al. (1983) Purified human basophils and mast cells: current concepts of mediator release. Eur J Respir Dis 64: 53-61. |
| [41] |
Lueke AJ, Meeusen JW, Donato LJ, et al. (2016) Analytical and clinical validation of an LC-MS/MS method for urine leukotriene E4: a marker of systemic mastocytosis. Clin Biochem 49: 979-982. https://doi.org/10.1016/j.clinbiochem.2016.02.007
|
| [42] |
Wenzel SE, Trudeau JB, Kaminsky DA, et al. (1995) Effect of 5-lipoxygenase inhibition on bronchoconstriction and airway inflammation in nocturnal asthma. Am J Resp Crit Care 152: 897-905. https://doi.org/10.1164/ajrccm.152.3.7663802
|
| [43] |
Mackfarlane AJ, Dworski R, Sheller JR, et al. (2000) Sputum cysteinyl leukotrienes increase 24 hours after allergen inhalation in atopic asthmatics. Am J Resp Crit Care 161: 1553-1558. https://doi.org/10.1164/ajrccm.161.5.9906068
|
| [44] |
Motomura C, Ide K, Shimoda T, et al. (2021) Exercise-induced anaphylaxis unrelated to food ingestion and with hyperleukotrieneuria during challenge testing. Allergy Asthma Clin Immunol 17: 89-94. https://doi.org/10.1186/s13223-021-00593-8
|
| [45] | Raithel M, Zopf Y, Kimpel S, et al. (2011) The measurement of leukotrienes in urine as diagnostic option in systemic mastocytosis. J Physiol Pharmacol 62: 469-472. |
| [46] |
Butterfield JH (2010) Increased leukotriene E4 excretion in systemic mastocytosis. Prostag Oth Lipid M 92: 73-76. https://doi.org/10.1016/j.prostaglandins.2010.03.003
|

Figures(2)

Peter Jandus. Arachidonic acid metabolism and its use in the diagnosis of mastocytosis[J]. AIMS Allergy and Immunology, 2023, 7(1): 82-91. doi: 10.3934/Allergy.2023006







 DownLoad:
DownLoad: