Citation: Rocio Caja Rivera, Shakir Bilal, Edwin Michael. The relation between host competence and vector-feeding preference in a multi-host model: Chagas and Cutaneous Leishmaniasis[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5561-5583. doi: 10.3934/mbe.2020299

| [1] | D. Biswas, S. Dolai, C. Jahangir, P. K. Roy, E. V. Grigorieva, Cost-Effective Analysis of Control Strategies to Reduce the Prevalence of Cutaneous Leishmaniasis, Based on a Mathematical Model, Math. Comput. Appl., 23 (2018), 38. |
| [2] | R. Caja Rivera, I. Barradas, Vector Preference Annihilates Backward Bifurcation and Reduces Endemicity, Bull. Math. Biol., 81 (2019), 4447-4469. |
| [3] | S. S. Gervasi, D. J. Civitello, H. J. Kilvitis, L. B. Martin, The context of host competence: a role for plasticity in host-parasite dynamics, Trends Parasitol., 31 (2015), 419-425. |
| [4] |
J. E. Simpson, P. J. Hurtado, J. Medlock, G. Molaei, T. G. Andreadis, A. P. Galvani, et al., Vector host-feeding preferences drive transmission of multi-host pathogens: West Nile virus as a model system, Proc. Royal Soc. B, 279 (2012), 925-933. doi: 10.1098/rspb.2011.1282

|
| [5] | L. Yakob, M. B. Bonsall, G. Yan, Modelling knowlesi malaria transmission in humans: vector preference and host competence, Malaria J., 9 (2010), 329. |
| [6] | J. E. Rabinovich, O. Rossell, Mathematical models and ecology of Chagas disease, American Trypanosomiasis Research, PAHO Sci. Publ, 318 (1976), 245-250. |
| [7] | J. E. Cohen, R. E. Gürtler, Modeling household transmission of American trypanosomiasis, Science, 293 (2001), 694-698. |
| [8] | C. S. Apperson, K. Hassan, B. A. Harrison, H. M. Savage, S. E. Aspen, A. Farajollahi, et al., Host feeding patterns of established and potential mosquito vectors of West Nile virus in the eastern United States, Vector-Borne Zoonotic Dis., 4 (2004), 71-82. |
| [9] |
F. Keesing, M. H. Hersh, M. Tibbetts, D. J. McHenry, S. Duerr, J. Brunner, et.al., Reservoir competence of vertebrate hosts for Anaplasma phagocytophilum, Emerging Infect. Dis., 18 (2012), 2013. doi: 10.3201/eid1812.120919
|
| [10] | C. L. Hodo, S. A. Hamer, Toward an ecological framework for assessing reservoirs of vector-borne pathogens: wildlife reservoirs of Trypanosoma cruzi across the southern United States, ILAR J., 58 (2017), 379-392. |
| [11] | R. S. Ostfeld, F. Keesing, Biodiversity series: the function of biodiversity in the ecology of vectorborne zoonotic diseases, Can. J. Zool., 78 (2000), 2061-2078. |
| [12] |
T. Lembo, K. Hampson, D. T. Haydon, M. Craft, A. Dobson, J. Dushoff, et al., Exploring reservoir dynamics: a case study of rabies in the Serengeti ecosystem, J. Appl. Ecol., 45 (2008), 1246-1257. doi: 10.1111/j.1365-2664.2008.01468.x
|
| [13] | P. J. Hudson, A. P. Rizzoli, B. T. Grenfell, J. A. P. Heesterbeek, A. P. Dobson, Ecology of wildlife diseases, 2002, 1-5. |
| [14] | R. E. Gürtler, M. V. Cardinal, Reservoir host competence and the role of domestic and commensal hosts in the transmission of Trypanosoma cruzi, Acta Trop., 151 (2015), 32-50. |
| [15] | D. Richter, A. Spielman, N. Komar, F. R. Matuschka, Competence of American robins as reservoir hosts for Lyme disease spirochetes, Emerging Infect. Dis., 6 (2000), 133. |
| [16] | J. L. Brunner, K. LoGiudice, R. S. Ostfeld, Estimating reservoir competence of Borrelia burgdorferi hosts: prevalence and infectivity, sensitivity, and specificity, J. Med. Entomol., 45 (2008), 139-147. |
| [17] | F. Keesing, R. D. Holt, R. S. Ostfeld, Effects of species diversity on disease risk, Ecol. Lett., 9 (2006), 485-498. |
| [18] | L. Yakob, How do biting disease vectors behaviourally respond to host availability?, Parasite. Vector., 9 (2016), 468. |
| [19] | A. M. Kilpatrick, L. D. Kramer, M. J. Jones, P. P. Marra, P. Daszak, West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior, PLoS Biol., 4 (2006), e82. |
| [20] | W. Takken, N. O. Verhulst, Host preferences of blood-feeding mosquitoes, Annu. Rev. Entomol., 58 (2013), 433-453. |
| [21] | P. Queiroz, G. Monteiro, V. Macedo, M. Rocha, L. Batista, J. Queiroz, et al., Canine visceral leishmaniasis in urban and rural areas of Northeast Brazil, Res. Vet. Sci., 86 (2009), 267-273. |
| [22] | R. Gürtler, E. Ricardo, L. A. Ceballos, P. Ordóñez-Krasnowski, L. A. Lanati, R. Stariolo, et al., Strong host-feeding preferences of the vector Triatoma infestans modified by vector density: implications for the epidemiology of Chagas disease, PLoS Negl. Trop. Dis., 3 (2009), e447. |
| [23] | H. V. Pates, W. Takken, K. Stuke, C. F. Curtis, Differential behaviour of Anopheles gambiae sensu stricto (Diptera: Culicidae) to human and cow odours in the laboratory, Bull. Entomol. Res., 91 (2001), 289-296. |
| [24] | D. T. Haydon, S. Cleaveland, L. H. Taylor, M. Karen Laurenson, Identifying reservoirs of infection: a conceptual and practical challenge, Emerging Infect. Dis., 8 (2002), 1468-1473. |
| [25] | R. Ostfeld, F. Keesing, Biodiversity series: the function of biodiversity in the ecology of vectorborne zoonotic diseases, Canadian J. Zool., 78 (2000), 2061-2078. |
| [26] | E. Miller, A. Huppert, The effects of host diversity on vector-borne disease: the conditions under which diversity will amplify or dilute the disease risk, PLoS One, 8 (2013), e80279. |
| [27] | R. Ostfeld, F. Keesing, V. T. Eviner, Infectious disease ecology: effects of ecosystems on disease and of disease on ecosystems, Princeton University Press, 2008. |
| [28] | R. M. Anderson, R. M. Robert, Infectious diseases of humans: dynamics and control, Oxford University Press, 1992. |
| [29] | P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. |
| [30] | W. B. Karesh, A. Dobson, J. O. Lloyd-Smith, J. Lubroth, M. A. Dixon, M. Bennett, et al., Ecology of zoonoses: natural and unnatural histories, The Lancet, 380 (2012), 1936-1945. |
| [31] | Y. Hashiguchi, E. Gomez, A. G. Cáceres, L. N. Velez, N. V. Villegas, K. Hashiguchi, et al., Andean cutaneous leishmaniasis (Andean-CL, uta) in Peru and Ecuador: the causative Leishmania parasites and clinico-epidemiological features, Acta Trop., 177 (2018), 135-145. |
| [32] | E. A. Llanos-Cuentas, N. Roncal, P. Villaseca, L. Paz, E. Ogusuku, J. E. Perez, et al., Natural infections of Leishmania peruviana in animals in the Peruvian Andes, T. Roy. Soc. Trop. Med. H., 93 (1999), 15-20. |
| [33] | J. Arevalo, L. Ramirez, V. Adaui, M. Zimic, G. Tulliano, C. Miranda-Verástegui, et al., Influence of Leishmania (Viannia) species on the response to antimonial treatment in patients with American tegumentary leishmaniasis, J. Infect. Dis., 195 (2007), 1846-1851. |
| [34] | Y. Hashiguchi, A. G. Cáceres, L. N. Velez, N. V. Villegas, K. Hashiguchi, T. Mimori, et al., Andean cutaneous leishmaniasis (Andean-CL, uta) in Peru and Ecuador: the vector Lutzomyia sand flies and reservoir mammals, Acta Trop., 178 (2018), 264-275. |
| [35] | Y. Hashiguchi, E. A. Gomez, V. V. De Coronel, T. Mimori, M. Kawabata, M. Furuya, et al., Andean leishmaniasis in Ecuador caused by infection with Leishmania mexicana and L. majorlike parasites, Am. J. Trop. Med. H., 44 (1991), 205-217. |
| [36] | R. Reithinger, C. R. Davies, Is the domestic dog (Canis familiaris) a reservoir host of American cutaneous leishmaniasis? A critical review of the current evidence, Am. J. Trop. Med. H., 61 (1999), 530-541. |
| [37] | C. R. Davies, E. A. Llanos-Cuentas, P. Campos, J. Monge, P. Villaseca, C. Dye, Cutaneous leishmaniasis in the Peruvian Andes: risk factors identified from a village cohort study, Am. J. Trop. Med. H., 56 (1997), 85-95. |
| [38] | E. A. Llanos-Cuentas, C. Davies, Epidemiological studies on Andean Cutaneous Leishmaniasis and their significance for designing a control strategy, In Leishmaniasis Control Strategies: a Critical Evaluation of IDRC Supported Research; proceedings of a workshop held in Mérida, Mexico, Nov. 25-29, 1991... IDRC, Ottawa, ON, CA, 1992. |
| [39] | I. Barradas, R. M. Caja Rivera, Cutaneous leishmaniasis in Peru using a vector-host model: Backward bifurcation and sensitivity analysis, Math. Methods Appl. Sci., 41 (2018), 1908-1924. |
| [40] | J. E. Rabinovich, M. D. Feliciangeli, Parameters of Leishmania braziliensis transmission by indoor Lutzomyia ovallesi in Venezuela, Am. J. Trop. Med. H., 70 (2004), 373-382. |
| [41] | S. L. Sánchez, A. E. Sáenz, M. J. Pancorbo, D. R. Zegarra, V. N. Garcés, R. A. Regis, Leishmaniasis: Dermatología, 14 (2004), 82-98. |
| [42] | CDC: Center for Disease Control and Prevention, available from: https://www.cdc.gov/parasites/chagas/. |
| [43] | J. R. Coura, Chagas disease:control, elimination and eradication, Is it possible?, Mem. Inst. Oswaldo Cruz, 108 (2013), 962-967. |
| [44] | M. P. Barrett, S. L. Croft, Management of trypanosomiasis and leishmaniasis, Brit. Med. Bull., 104 (2012), 175-196. |
| [45] | A. L. Roque, A. M. Jansen, Wild and synanthropic reservoirs of Leishmania species in the Americas, Int. J. Parasitol-Par., 3 (2014), 251-262. |
| [46] | L. F. Chaves, M. J. Hernandez, S. Ramos, Simulación de modelos matemáticos como herramienta para el estudio de los reservorios de la Leishmaniasis Cutánea Americana, Divulgaciones Matemáticas, 16 (2008), 125-154. |
| [47] | J. E. Rabinovich, C. Wisnivesky-Colli, N. D. Solarz, R. E. Gürtler, Probability of transmission of Chagas disease by Triatoma infestans (Hemiptera: Reduviidae) in an endemic area of Santiago del Estero, Argentina, Bull. World Health Organ., 68 (1990), 737. |
| [48] | C. Kribs-Zaleta, Estimating contact process saturation in sylvatic transmission of Trypanosoma cruzi in the United States, PLoS Negl. Trop. Dis., 4 (2010), e656. |
| [49] | Pan American Health Organization-World Health Organization. |
| [50] | C. Pirmez, S. G. Coutinho, M. C. Marzochi, M. P. Nunes, G. Grimaldi, Canine American cutaneous leishmaniasis: a clinical and immunological study in dogs naturally infected with Leishmania braziliensis braziliensis in an endemic area of Rio de Janeiro, Brazil, Am. J. Trop. Med. H., 38 (1988), 52-58. |
| [51] | L. Reveiz, A. N. Maia-Elkhoury, R. S. Nicholls, G. A. Sierra Romero, Z. E. Yadon, Interventions for American cutaneous and mucocutaneous leishmaniasis: a systematic review update, PloS One, 8 (2013), e61843. |
| [52] | R. E. Gürtler, M. C. Cecere, M. A. Lauricella, M. V. Cardinal, U. Kitron, J. E. Cohen, Domestic dogs and cats as sources of Trypanosoma cruzi infection in rural northwestern Argentina, Parasitology, 134 (2007), 69-82. |
| [53] | M. B. Castañera, J. P. Aparicio, R. E. Gürtler, A stage-structured stochastic model of the population dynamics of Triatoma infestans, the main vector of Chagas disease, Ecol. Model., 162 (2003), 33- 53. |
| [54] | Ministerio de Salud y Protección Social (MinSalud), Enfermedad de Chagas. Memorias.p.1-34. |
| [55] | S. A. Pedro, H. E. Tonnang, S. Abelman, Uncertainty and sensitivity analysis of a Rift Valley fever model, Appl. Math. Comput., 279 (2016), 170-186. |
| [56] | C. Costa, R. Gomes, M. Silva, L. M. Garcez, P. Ramos, R. S. Santos, et al., Competence of the human host as a reservoir for Leishmania chagasi, J. Infect. Dis., 182 (2000), 997-1000. |
| [57] | J. R. Lockwood III, On the statistical analysis of multiple-choice feeding preference experiments, Oecologia, 116 (1998), 475-481. |
| [58] | T. D. Hollingsworth, E. R. Adams, R. M. Anderson, K. Atkins, S. Bartsch, M.G. Basáñez, et al., Quantitative analyses and modelling to support achievement of the 2020 goals for nine neglected tropical diseases, Parasite. Vector., 8 (2015), 630. |
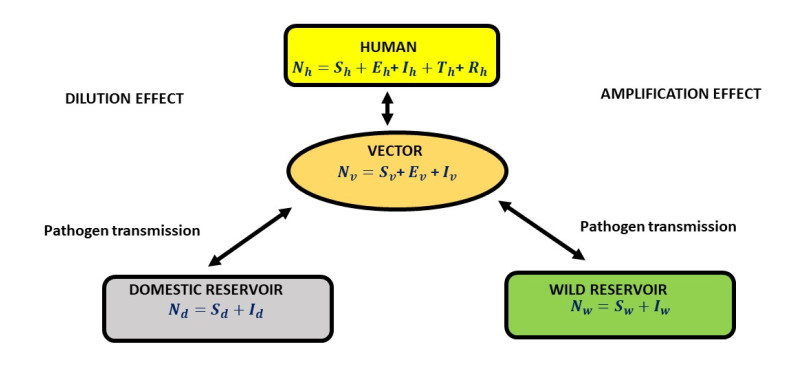
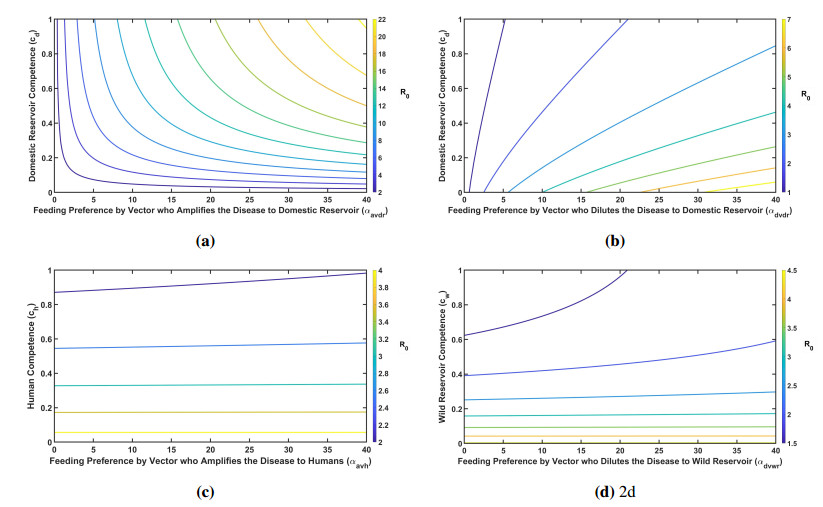
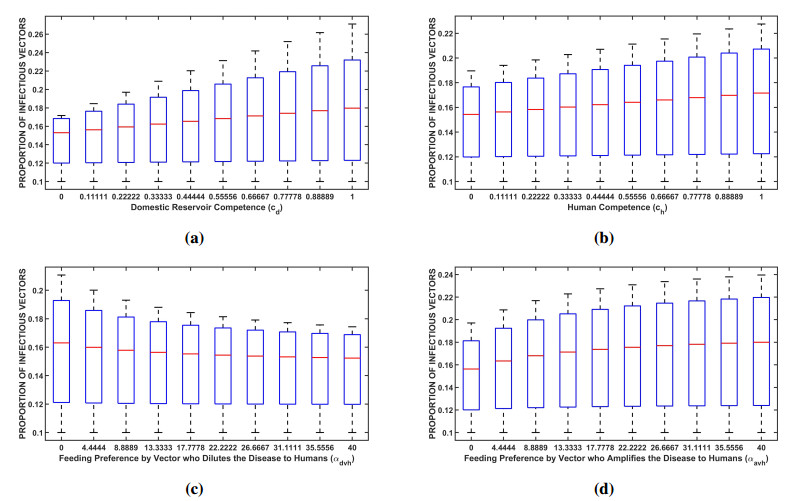
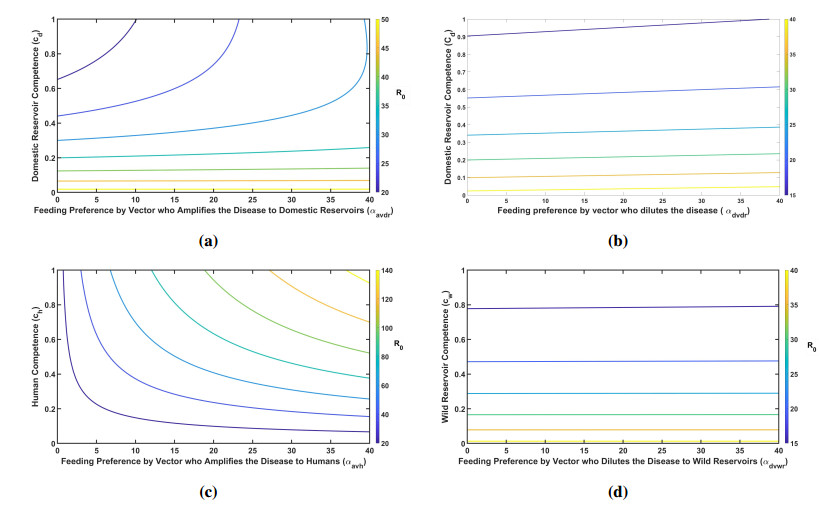
Figures(7) / Tables(3)

Rocio Caja Rivera, Shakir Bilal, Edwin Michael. The relation between host competence and vector-feeding preference in a multi-host model: Chagas and Cutaneous Leishmaniasis[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5561-5583. doi: 10.3934/mbe.2020299







 DownLoad:
DownLoad: