Citation: Saswati Biswas, Pankaj Kumar Tiwari, Yun Kang, Samares Pal. Effects of zooplankton selectivity on phytoplankton in an ecosystem affected by free-viruses and environmental toxins[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1272-1317. doi: 10.3934/mbe.2020065

| [1] | R. M. Anderson and R. M. May, The invasion, persistence, and spread of infectious diseases within animal and plant communities, Philos. Trans. R. Soc. Lond. B, 314 (1986), 533-570. |
| [2] | J. Chattopadhyay and N. Bairagi, Pelicans at risk in Salton Sea - an eco-epidemiological model, Ecol. Model., 136 (2001), 103-112. |
| [3] | Y. Xiao and L. Chen, Modelling and analysis of a predator-prey model with disease in the prey, Math. Biosci., 171 (2001), 59-82. |
| [4] | E. Venturino, Epidemics in predator-prey models: Disease in the predators, IMA J. Math. Appl. Med. Biol., 19 (2002), 185-205. |
| [5] | K. Hadeler and H.I. Freedman, Predator-prey population with parasite infection, J. Math. Biol., 27 (1989), 609-631. |
| [6] | S. Samanta, R. Dhar, J. Pal, et al., Effect of enrichment on plankton dynamics where phytoplankton can be infected from free viruses, Nonlinear Studies, 20 (2013), 223-236. |
| [7] | K. D. Lafferty and A. K. Morris, Altered behaviour of parasitized killfish increases susceptibility to predation by bird final hosts, Ecology, 77 (1996), 1390-1397. |
| [8] | W. R. DeMott, Optimal foraging theory as a predictor of chemically mediated food selection by suspension-feeding copepods, Limnol. Oceanogr., 34 (1989), 140-154. |
| [9] | M. A. Leibold, Biodiversity and nutrient enrichment in pond plankton communities, Evol. Ecol. Res., 1 (1999), 73-95. |
| [10] | J. L. Brooks and S. I. Dodson, Predation, body size, and composition of plankton, Science, 150 (1965), 28-35. |
| [11] | A. Mitra, C. Castellanib, W. C. Gentleman, et al., Bridging the gap between marine biogeochemical and fisheries sciences; configuring the zooplankton link, Prog. Oceanogr., 129 (2014), 176-199. |
| [12] | M. D. Troch, M. Grego, V. A. Chepurnov, et al., Food patch size, food concentration and grazing efficiency of the harpacticoid Paramphiascella fulvofasciata (Crustacea, Copepoda), J. Exp. Mar. Biol. Ecol., 343 (2007), 210-216. |
| [13] | W. R. DeMott, Discrimination between algae and artificial particles by freshwater and marine copepods, Limnol. Oceanogr., 33 (1988), 397-408. |
| [14] | J. Pal, S. Bhattacharya and J. Chattopadhyay, Does predator go for size selection or preferential toxic-nontoxic species under limited resource?, OJBS, 10 (2010), 11-16. |
| [15] | N. Aberle, H. Hillebrand, J. Grey, et al., Selectivity and competitive interactions between two benthic invertebrate grazers (Asellus aquaticus and Potamopyrgus antipodarum): An experimental study using 13C-and 15N-labelled diatoms, Freshwater Biol., 50 (2005), 369-379. |
| [16] | M. G. Danielsdottir, M. T. Brett, G. B. Arhonditsis, Phytoplankton food quality control of planktonic food web processes, Hydrobiologia, 589 (2007), 29-41. |
| [17] |
M. Huntley, P. Sykes, S. Rohan, et al., Chemically-mediated rejection of dinoflagellate prey by the copepods Calanus pacificus and Paracalanus parvus: mechanism, occurrence and significance, Mar. Ecol. Prog. Ser., 28 (1986), 105-120. doi: 10.3354/meps028105

|
| [18] | R. S. Fulton III and H.W. Paerl, Effects of colonial morphology on zooplankton utilization of algal resources during blue-green algal (Microcystis aeruginosa) blooms, Limnol. Oceanogr. 32 (1987), 634-644. |
| [19] | G. A. Paffenhofer and K. B. Van Sant, The feeding response of a marine planktonic copepod to quantity and quality of particles, Mar. Ecol. Prog. Ser. 27 (1985), 55-65. |
| [20] | C. Evans, D. W. Pond and W. H. Wilson, Changes in Emiliania huxleyi fatty acid profiles during infection with E. huxleyi virus 86: physiological and ecological implications, Aquat. Microb. Ecol., 55 (2009), 219-228. |
| [21] | G. Bratbak, J. K. Egge and M. Heldal, Viral mortality of the marine alga Emiliania huxleyi (Haptophyceae) and termination of algal blooms, Mar. Ecol. Prog. Ser., 93 (1993), 39-48. |
| [22] | C. Evans and W.H. Wilson, Preferential grazing of Oxyrrhis marina on virus infected Emiliania huxleyi, Limnol. Oceanogr., 53 (2008), 2035-2040. |
| [23] |
A. Vermont, J. M. Martnez, J. D. Waller, et al., Virus infection of Emiliania huxleyi deters grazing by the copepod Acartia tonsa, J. Plankton Res., 38 (2016), 1194-1205. doi: 10.1093/plankt/fbw064
|
| [24] |
D. W. Townsend, M. D. Keller, P. M. Holligan, et al., Blooms of the coccolithophore Emiliania huxleyi with respect to hydrography in the Gulf of Maine, Cont. Shelf Res., 14 (1994), 979-1000. doi: 10.1016/0278-4343(94)90060-4
|
| [25] | W. H. Wilson, G. A. Tarran, D. Schroeder, et al., Isolation of viruses responsible for the demise of an Emiliania huxleyi bloom in the English Channel, J. Mar. Biol. Assoc. U.K., 82 (2002), 369-377. |
| [26] | C. Evans, S. V. Kadner, L. J. Darroch, et al., The relative significance of viral lysis and microzooplankton grazing as pathways of dimethylsulfoniopropionate (DMSP) cleavage: An Emiliania huxleyi culture study, Limnol. Oceanogr., 52 (2007), 1036-1045. |
| [27] |
S. Strom, G. Wolfe, J. Holmes, et al., Chemical defense in the microplankton I: Feeding and growth rates of heterophic protists on the DMS-producing phytoplankter Emiliania huxleyi, Limnol. Oceanogr., 48 (2003), 217-229. doi: 10.4319/lo.2003.48.1.0217
|
| [28] | G. V. Wolfe and M. Steinke, Grazing-activated production of dimethyl sulfide (DMS) by two clones of Emiliania huxleyi, Limnol. Oceanogr., 41 (1996), 1151-1160. |
| [29] | M. Steinke, G. Malin and P. S. Liss, Trophic interaction in the sea: an ecological role for climate relevant volatiles? J. Phycol., 38 (2002), 630-630. |
| [30] | E. Beretta and Y. Kuang, Modeling and analysis of a marine bacteriophage infection, Math. Biosci., 149 (1998), 57-76. |
| [31] | E. Beltrami and T.O. Carroll, Modeling the role of viral disease in recurrent phytoplankton blooms, J. Math. Biol., 32 (1994), 857-863. |
| [32] | S. Gakkhar and K. Negi, A mathematical model for viral infection in toxin producing phytoplankton and zooplankton system, Appl. Math. Comp., 179 (2006), 301-313. |
| [33] | J. Chattopadhyay and S. Pal, Viral infection on phytoplankton-zooplankton system: A mathematical model, Ecol. Model., 151 (2002), 15-28. |
| [34] |
N. Bairagi, P. K. Roy, R. R. Sarkar, et al., Virus replication factor may be a controlling agent for obtaining disease-free system in a multi-species eco-epidemiological system, J. Biol. Syst., 13 (2005), 245-259. doi: 10.1142/S0218339005001501
|
| [35] | J. Labille and J. Brant, Stability of nanoparticles in water, Nanomedicine, 5 (2010), 985-998. |
| [36] | A. J. Miao, K. A. Schwehr, C. Xu, et al., The algal toxicity of silver engineered nanoparticles and detoxification by exopolymeric substances, Environ. Pollut., 157 (2009), 3034-3041. |
| [37] | R. J. Miller, S. Bennett, A. A. Keller, et al., TiO2 nanoparticles are phototoxic to marine phytoplankton, PLoS ONE, 7 (2012), e30321. |
| [38] | S. Rana, S. Samanta, S.Bhattacharya, et al., The effect of nanoparticles on plankton dynamics: A mathematical model, BioSystems, 127 (2015), 28-41. |
| [39] | P. Panja, S. K. Mondal and D. K. Jana, Effects of toxicants on phytoplankton-zooplankton-fish dynamics and harvesting, Chaos Solit. Fract., 104 (2017), 389-399. |
| [40] | X. Yu, S. Yuan and T. Zhang, Survival and ergodicity of a stochastic phytoplankton-zooplankton model with toxin-producing phytoplankton in an impulsive polluted environment, Appl. Math. Comp., 347 (2019), 249-264. |
| [41] | N. Bairagi and D. Adak, Complex dynamics of a predator-prey-parasite system: An interplay among infection rate, predator's reproductive gain and preference, Ecol. Compl., 22 (2015), 1-12. |
| [42] | N. Bairagi, S. Saha, S. Chaudhuri, et al., Zooplankton selectivity and nutritional value of phytoplankton influences a rich variety of dynamics in a plankton population model, Phy. Rev. E, 99 (2019), 012406. |
| [43] | K. Bester, H. Hhnerfuss, U. Brockmann, et al., Biological effects of triazine herbicide contamination on marine phytoplankton, Arch. Environ. Contam. Toxicol., 29 (1995), 277-283. |
| [44] | R. C. Antweiler, C. J. Patton and H. E. Taylor, Nutrients, in chemical data for water samples collected during four upriver cruises on the Mississippi river between New Orleans, Louisiana, and Minneapolis, Minnesota, May 1990-April 1992, J.A. Moody, ed., U.S. Geological Survey Open-File Report, 94-523 (1995), 89-125. |
| [45] | U.S. Environmental Protection Agency Great Lakes National Program Office Significant Activities Report. http://www.epa.gov/glnpo/aoc/waukegan.html. |
| [46] | J. R. Rueter, S. W. Chisholm and F. Morel, Effects of copper toxicity on silicon acid uptake and growth in Thalassiosira pseudonana, J. Phycol. 17 (1981) 270-278. |
| [47] | J. C. Holmes and W. M. Bethel, Modification of intermediate host behavior by parasites, In: Canning, E.V., Wright, C.A. (Eds.), Behavioral Aspects of Parasite Transmission. Suppl. I to Zool. f. Linnean Soc., 51 (1972), 123-149. |
| [48] | K. D. Lafferty, Foraging on prey that are modified by parasites, Am. Nat., 140 (1992), 854-867. |
| [49] | W. D. Hamilton, R. Axelrod and R. Tanese, Sexual reproduction as an adaptation to resist parasite: a review, Proc. Natl. Acad. Sci. USA, 87 (1990), 3566-3573. |
| [50] | H. L. Smith, The Rosenzweig-Macarthur predator-prey model, https://math.la.asu.edu/halsmith/Rosenzweig.pdf. |
| [51] | S. M. Blower and H. Dowlatabadi, Sensitivity and uncertainty analysis of complex models of disease transmission: An HIV model, as an example, Int. Stat. Rev., 62 (1994), 229-243. |
| [52] | S. Marino, I. B. Hogue, C. J. Ray, et al., A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178-196. |
| [53] | M. L. Rosenzweig, Paradox of enrichment: destabilization of exploitation ecosystems in ecological time, Science, 171 (1971), 385. |
| [54] | A. C. Baudoux, A. Noordeloos, M. Veldhuis, et al., Virally induced mortality of Phaeocystis globosa during two spring blooms in temperate coastal waters, Aquat. Microb. Ecol., 44 (2006), 207-217. |
| [55] | C. Evans, S. D. Archer, J. Stphan, et al., Direct estimates of the contribution of viral lysis and microzooplankton grazing to the decline of a Micromonas spp. population, Aquat. Microb. Ecol., 30 (2003), 207-219. |
| [56] | A. Chhater, H. Purohit, R. Shanker, et al., Bacterial consortia for crude oil spill remediation, Wat. Sci. Technol., 34 (1996), 187-193. |
| [57] | B. D. Duval, UV from sunlight excites nanoparticles to kill phytoplankton in lab setting, (2012) http://earthsky.org/human-world/uv-from-sunlight-excites-nanoparticles-to-kill-phytoplankton-in-lab-setting/. |
| [58] | A. Miao, X. Y. Zhang, Z. Luo, et al., Zinc oxide-engineered nanoparticles: dissolution and toxicity to marine phytoplankton, Environ. Toxicol. Chem., 29 (2010), 2814-2822. |
| [59] | G. Birkhoff and G. C. Rota, Ordinary Differential Equations, 4th edn. John Wiley & Sons, New York (1989). |
| [60] | J. K. Hale, Introduction to Functional Differential Equations, vol. 99, Springer, Berlin (1993). |
| [61] | X. Yang, L. Chen and J. Chen, Permanence and positive periodic solution for the single-species nonautonomous delay diffusive models, Comput. Math. Appl., 32 (1996), 109-116. |
| [62] | V. Lakshmikantham, S. Leela and A. A. Martynyuk, Stability Analysis of Nonlinear Systems, Marcel Dekker, Inc., New York/Basel (1989). |
| [63] | G. Gandolfo, Economic Dynamics, Springer, New York, USA (1996). |
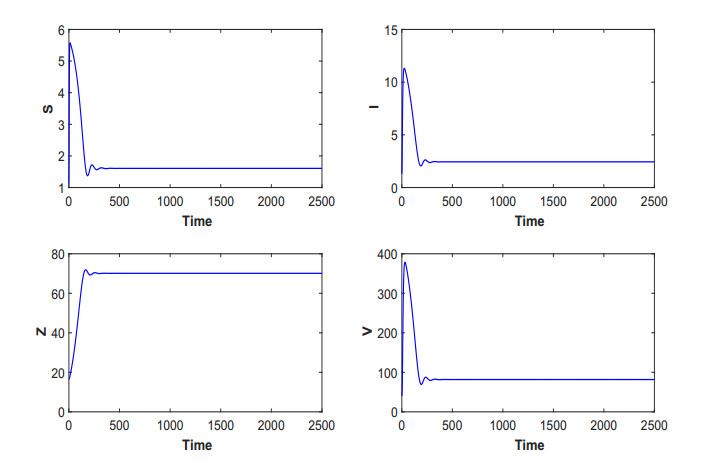
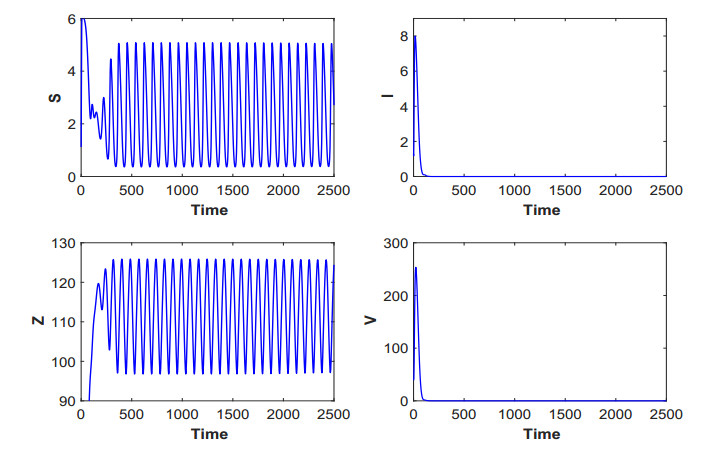
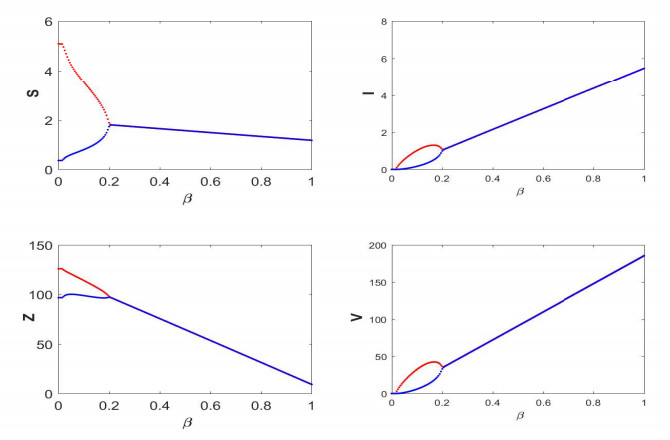
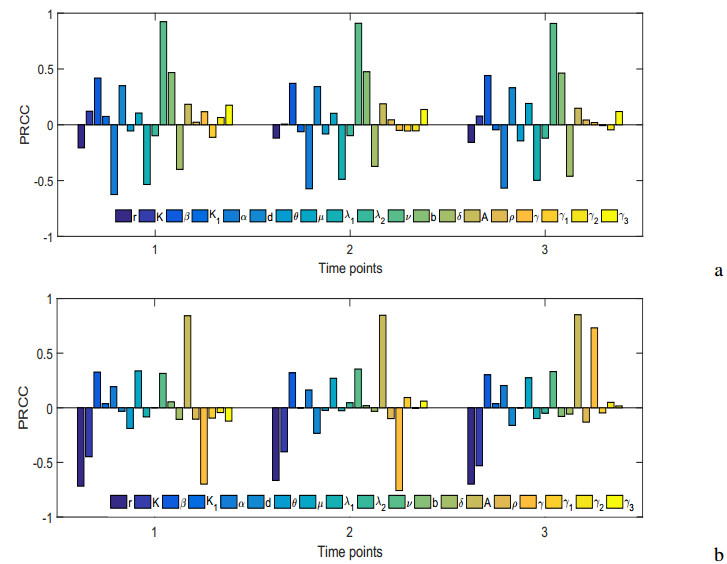
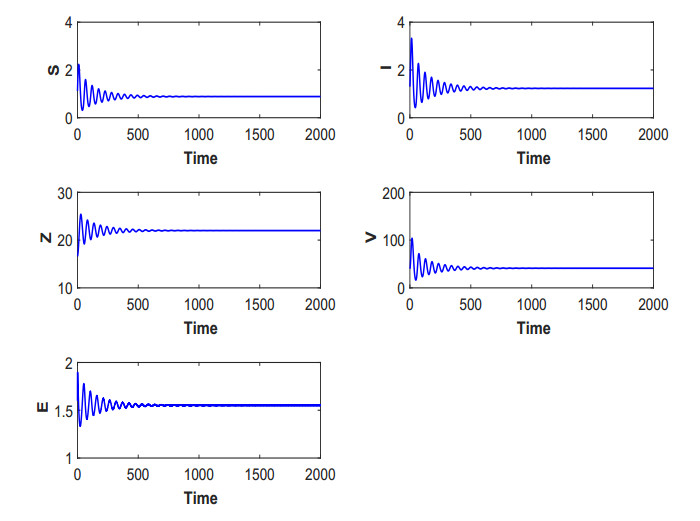
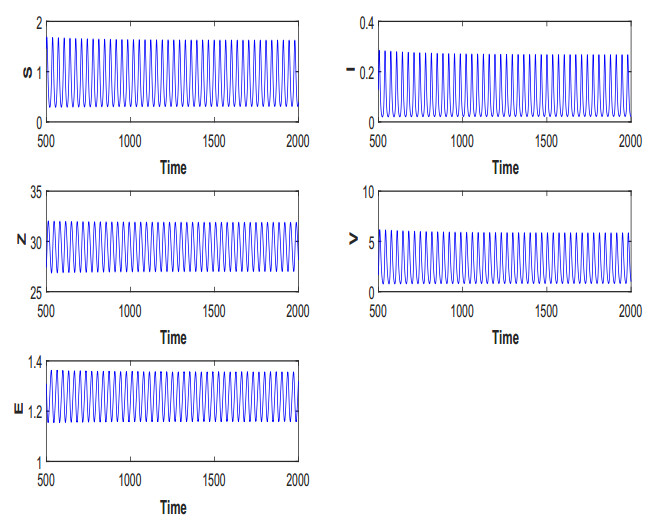
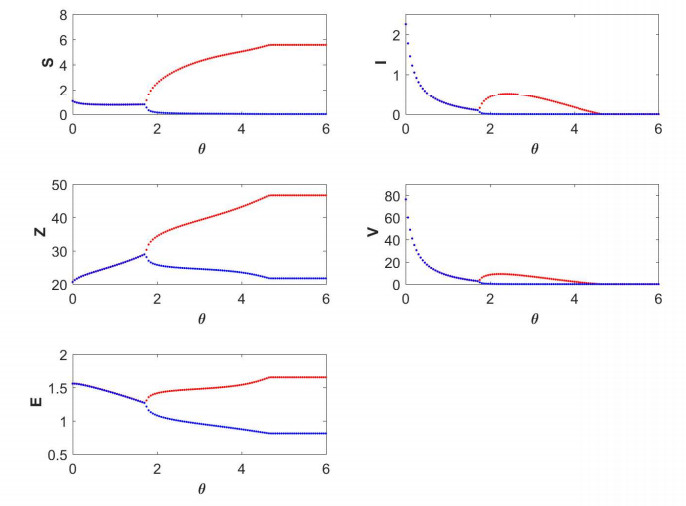
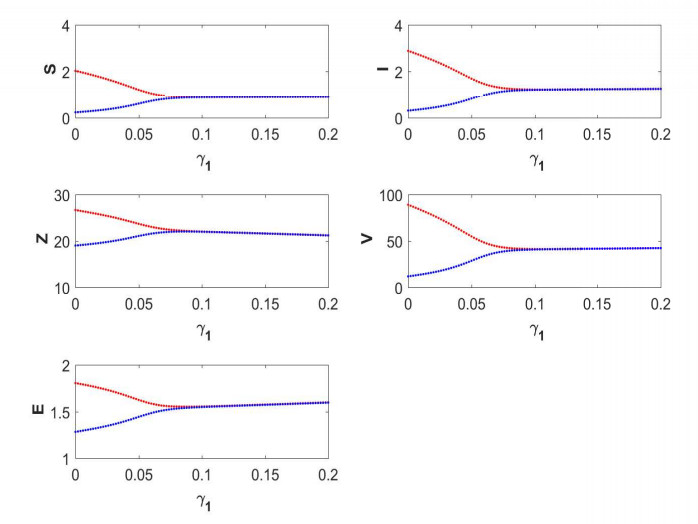
Figures(19) / Tables(5)

Saswati Biswas, Pankaj Kumar Tiwari, Yun Kang, Samares Pal. Effects of zooplankton selectivity on phytoplankton in an ecosystem affected by free-viruses and environmental toxins[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1272-1317. doi: 10.3934/mbe.2020065







 DownLoad:
DownLoad: