Cartilage tissue is avascular and resistant to tumor invasion, but the basis for these properties is still unclear. Here we report that the NH2-propeptide of type IIB procollagen (PIIBNP), a product of collagen biosynthesis, is capable of inhibiting angiogenesis both in vitro and in vivo. PIIBNP inhibits tube formation in human umbilical vein cells (HUVEC), inhibits endogenous endothelial cell outgrowth in mouse aortic ring angiogenesis bioassay and is anti-angiogenic in the mouse cornea angiogenesis assay. As αVß3 and αVß5 integrins are expressed primarily in endothelial cells, cancer cells and osteoclasts, but not in normal chondrocytes and PIIBNP binds to cell surface integrin αVß3 and αVß5, we propose that natural occurring PIIBNP protects cartilage by targeting endothelial cells during chondrogenesis, thus inhibiting angiogenesis, and rendering the tissue avascular.
Citation: Zhepeng Wang, Aiwu Lu. Cartilage type IIB procollagen NH2-propeptide, PIIBNP, inhibits angiogenesis[J]. AIMS Molecular Science, 2021, 8(4): 291-300. doi: 10.3934/molsci.2021022
Cartilage tissue is avascular and resistant to tumor invasion, but the basis for these properties is still unclear. Here we report that the NH2-propeptide of type IIB procollagen (PIIBNP), a product of collagen biosynthesis, is capable of inhibiting angiogenesis both in vitro and in vivo. PIIBNP inhibits tube formation in human umbilical vein cells (HUVEC), inhibits endogenous endothelial cell outgrowth in mouse aortic ring angiogenesis bioassay and is anti-angiogenic in the mouse cornea angiogenesis assay. As αVß3 and αVß5 integrins are expressed primarily in endothelial cells, cancer cells and osteoclasts, but not in normal chondrocytes and PIIBNP binds to cell surface integrin αVß3 and αVß5, we propose that natural occurring PIIBNP protects cartilage by targeting endothelial cells during chondrogenesis, thus inhibiting angiogenesis, and rendering the tissue avascular.

| [1] |
Forkman Y (1989) Successful treatment of an angiogenic disease. N Engl J Med 320: 1211-1212. doi: 10.1056/NEJM198905043201811

|
| [2] |
Schumacher BL, Block JA, Schmid TM (1994) A novel proteoglycan synthesized and secreted by chondrocytes of the superficial zone of articular cartilage. Arch Biochem Biophys 311: 144-152. doi: 10.1006/abbi.1994.1219
|
| [3] |
Jiang Y, Tuan RS (2015) Origin and function of cartilage stem/progenitor cells in osteoarthritis. Nat Rev Rheumatol 11: 206-212. doi: 10.1038/nrrheum.2014.200
|
| [4] |
Nyberg P, Xie L, Kalluri R (2005) Endogenous inhibitors of angiogenesis. Cancer Res 65: 3967-3979. doi: 10.1158/0008-5472.CAN-04-2427
|
| [5] |
Ryan MC, Sandell LJ (1990) Differential expression of a cysteine-rich domain in the amino-terminal propeptide of type II (cartilage) procollagen by alternative splicing of mRNA. J Biol Chem 265: 10334-10339. doi: 10.1016/S0021-9258(18)86950-2
|
| [6] |
Sandell LJ, Morris N, Robbins JR (1991) Alternatively spliced type II procollagen mRNAs define distinct populations of cells during vertebral development: differential expression of the amino-propeptide. J Cell Biol 114: 1307-1319. doi: 10.1083/jcb.114.6.1307
|
| [7] |
Zhu Y, Oganesian A, Keene DR (1999) Type IIA procollagen containing the cysteine-rich amino propeptide is deposited in the extracellular matrix of prechondrogenic tissue and binds to TGF-beta1 and BMP-2. J Cell Biol 144: 1069-1080. doi: 10.1083/jcb.144.5.1069
|
| [8] |
Kim YS, Liotta LA, Kohn EC (1993) Cancer invasion and metastasis. Hosp Pract (Off Ed) 28: 92-96. doi: 10.1080/21548331.1993.11442792
|
| [9] |
Millauer B, Shaver LK, Plate KH (1994) Glioblastoma growth inhibited in vivo by a dominant-negative Flk-1 mutant. Nature 367: 576-579. doi: 10.1038/367576a0
|
| [10] |
Schwartz M, London A (2008) Glaucoma as a neuropathy amenable to neuroprotection and immune manipulation. Prog Brain Res 173: 375-384. doi: 10.1016/S0079-6123(08)01126-6
|
| [11] |
Schultz NM, Bhardwaj S, Barclay C (2021) Global Burden of Dry Age-Related Macular Degeneration: A Targeted Literature Review. Clin Ther 43: 1792-1818. doi: 10.1016/j.clinthera.2021.08.011
|
| [12] |
Sumariwalla PF, Cao Y, Wu HL (2003) The angiogenesis inhibitor protease-activated kringles 1-5 reduces the severity of murine collagen-induced arthritis. Arthritis Res Ther 5: R32-39. doi: 10.1186/ar608
|
| [13] |
Becker CM, D'Amato RJ (2007) Angiogenesis and antiangiogenic therapy in endometriosis. Microvasc Res 74: 121-130. doi: 10.1016/j.mvr.2007.04.008
|
| [14] |
Kirk S, Frank JA, Karlik S (2004) Angiogenesis in multiple sclerosis: is it good, bad or an epiphenomenon? J Neurol Sci 217: 125-130. doi: 10.1016/j.jns.2003.10.016
|
| [15] |
Vagnucci AH, Li WW (2003) Alzheimer's disease and angiogenesis. Lancet 361: 605-608. doi: 10.1016/S0140-6736(03)12521-4
|
| [16] |
Lijnen HR (2008) Angiogenesis and obesity. Cardiovasc Res 78: 286-293. doi: 10.1093/cvr/cvm007
|
| [17] |
Alghsi GC, Rüegg C (2006) Vascular integrins in tumor angiogenesis: mediators and therapeutic targets. Endothelium 13: 113-135. doi: 10.1080/10623320600698037
|
| [18] |
Kumar CC (2003) Integrin alpha v beta 3 as a therapeutic target for blocking tumor-induced angiogenesis. Curr Drug Targets 4: 123-131. doi: 10.2174/1389450033346830
|
| [19] |
Wang Z, Bryan J, Franz C, et al. (2010) Type IIB procollagen NH(2)-propeptide induces death of tumor cells via interaction with integrins alpha(V)beta(3) and alpha(V)beta(5). J Biol Chem 285: 20806-20817. doi: 10.1074/jbc.M110.118521
|
| [20] |
Javaherian K, Park SY, Pickl WF (2002) Laminin modulates morphogenic properties of the collagen XVIII endostatin domain. J Biol Chem 277: 45211-45218. doi: 10.1074/jbc.M206358200
|
| [21] | Nicosia RF, Ottinetti A (1990) Growth of microvessels in serum-free matrix culture of rat aorta. A quantitative assay of angiogenesis in vitro. Lab Invest 63: 115-122. |
| [22] |
Prockop DJ, Sieron AL, Li SW (1998) Procollagen N-proteinase and procollagen C-proteinase. Two unusual metalloproteinases that are essential for procollagen processing probably have important roles in development and cell signaling. Matrix Biol 16: 399-408. doi: 10.1016/S0945-053X(98)90013-0
|
| [23] |
Sandell LJ, Nalin A, Reife R (1994) Alternative splice form of type II procollagen mRNA (IIA) is predominant in skeletal precursors and non-cartilaginous tissues during early mouse development. Dev Dynamics 199: 129-140. doi: 10.1002/aja.1001990206
|
| [24] |
Rogers MA, Birsner AE, D'Amato RJ (2007) The mouse cornea micropocket angiogenesis assay. Nat Proc 2: 2545-2550. doi: 10.1038/nprot.2007.368
|
| [25] |
Tang DG, Diglio CA, Bazaz R (1995) Transcriptional activation of endothelial cell integrin αv by protein kinase C activator 12(S)-HETE. J Cell Sci 108: 2629-2644. doi: 10.1242/jcs.108.7.2629
|
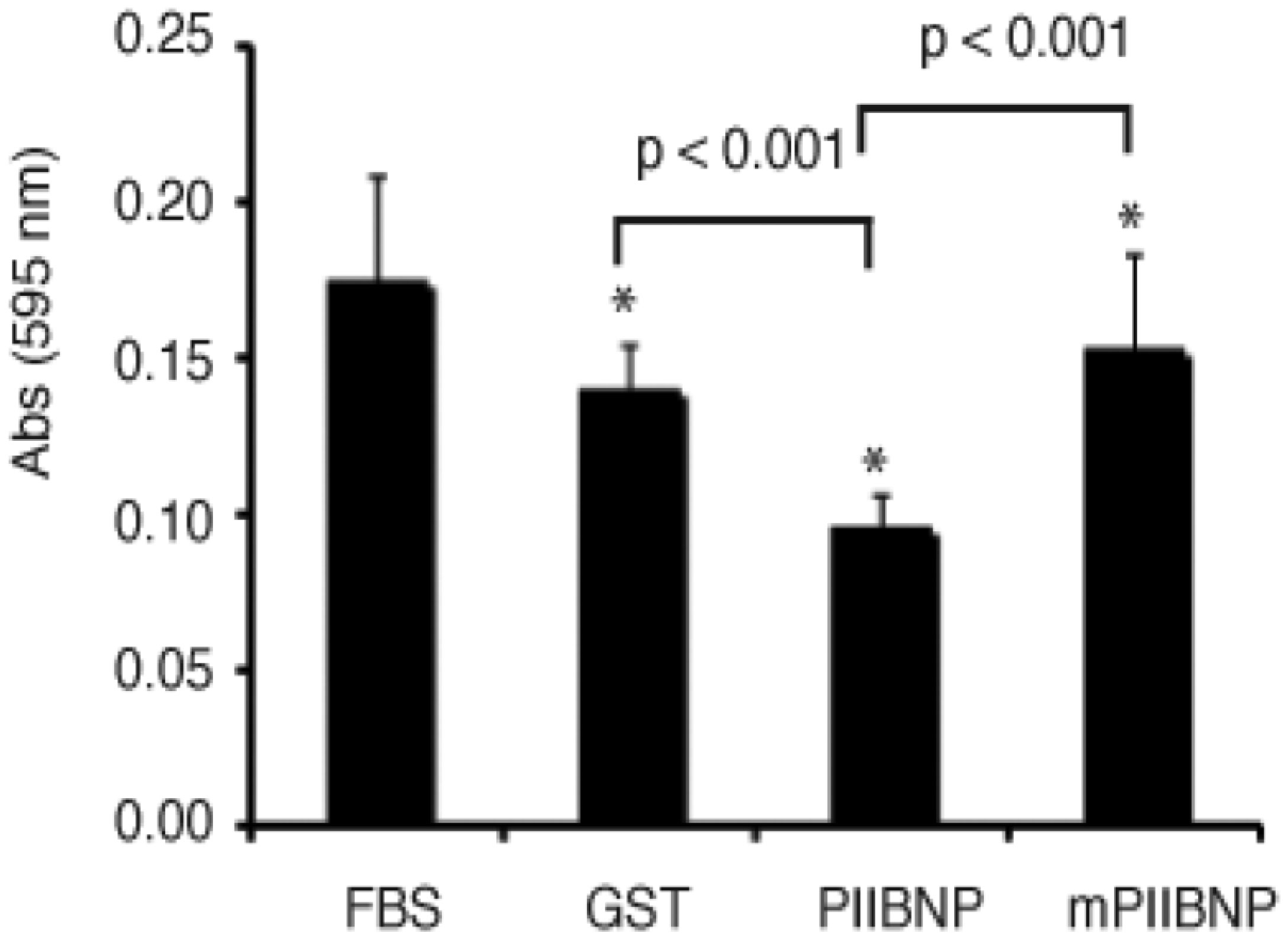
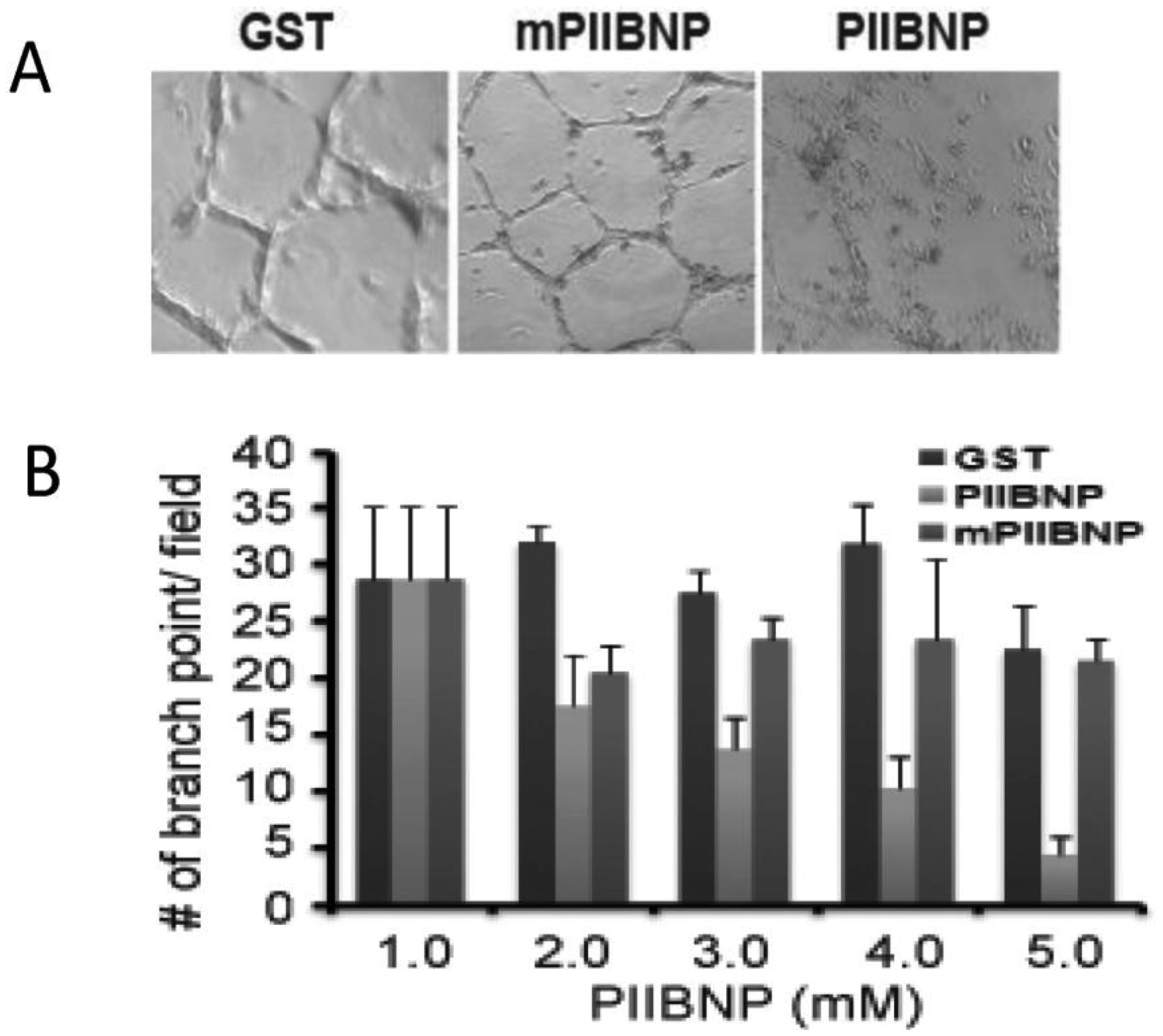
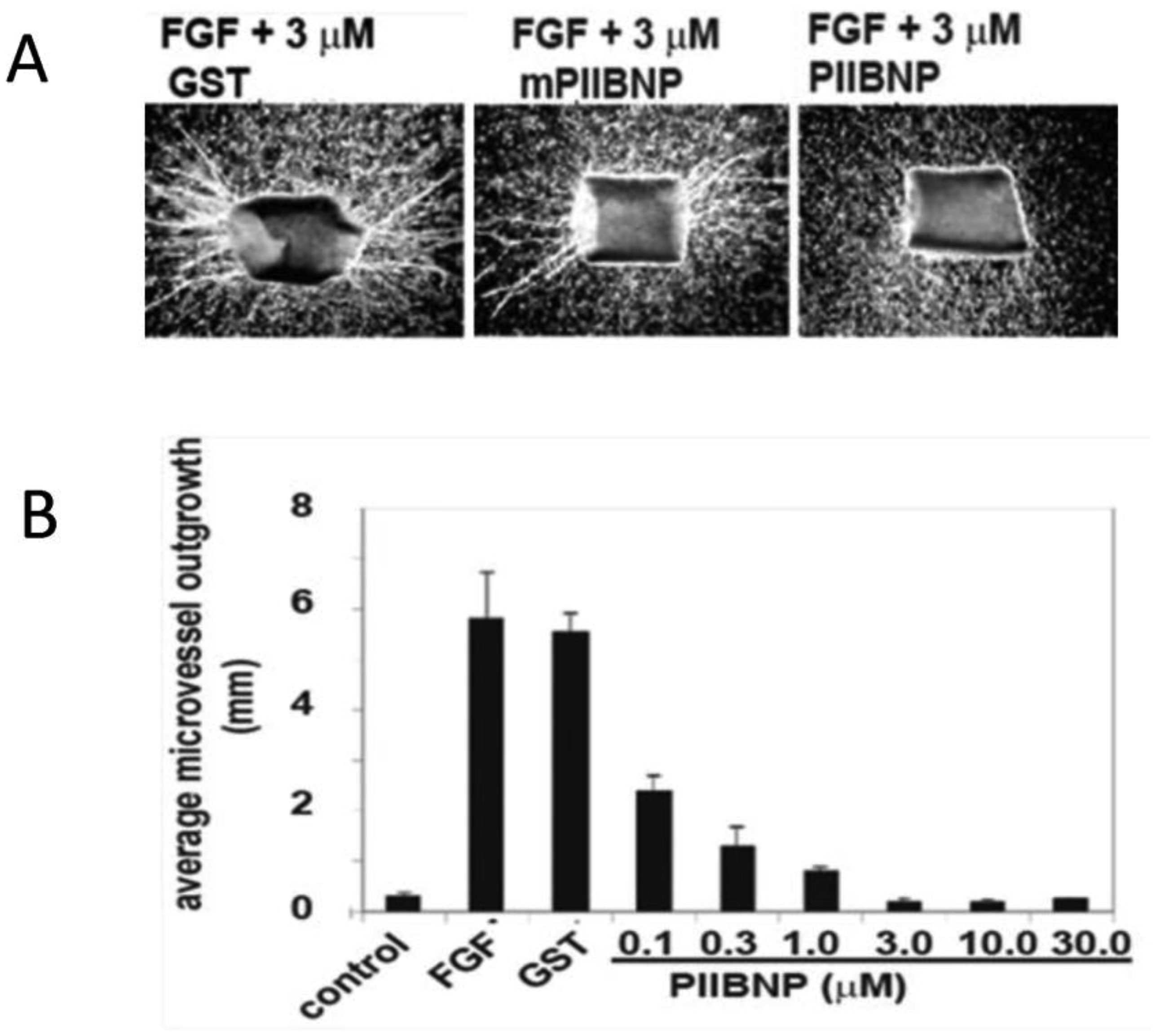
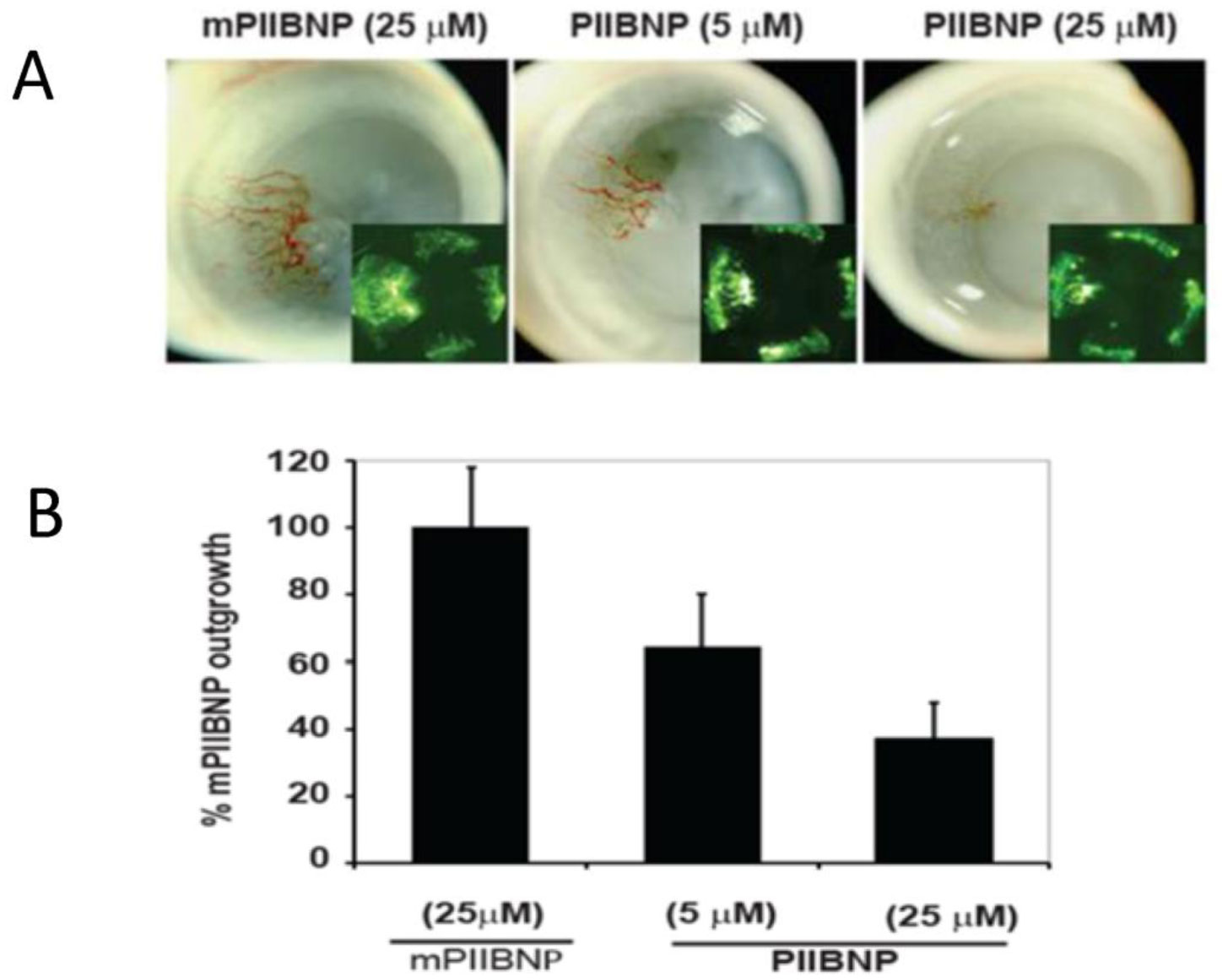
Figures(4)

Zhepeng Wang, Aiwu Lu. Cartilage type IIB procollagen NH2-propeptide, PIIBNP, inhibits angiogenesis[J]. AIMS Molecular Science, 2021, 8(4): 291-300. doi: 10.3934/molsci.2021022







 DownLoad:
DownLoad: