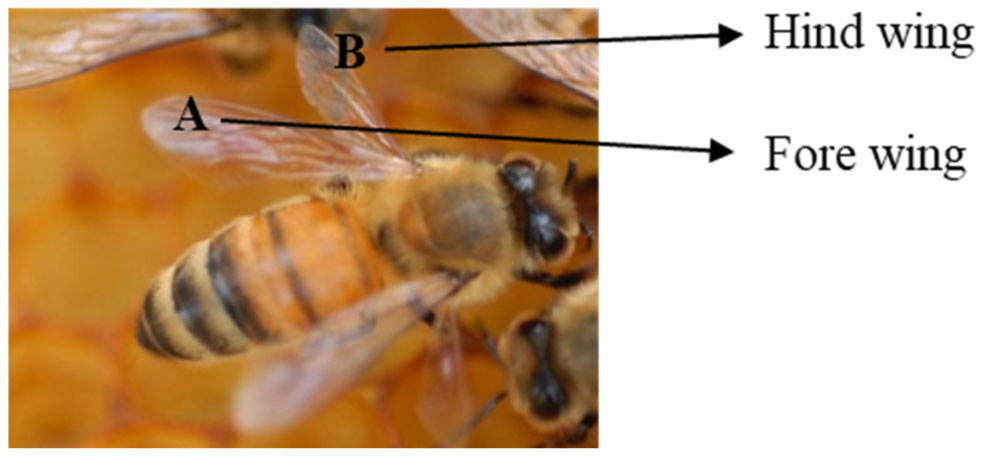
Natural surfaces with remarkable properties and functionality have become the focus of intense research. Heretofore, the natural antimicrobial properties of insect wings have inspired research into their applications. The wings of cicadas, butterflies, dragonflies, and damselflies have evolved phenomenal anti-biofouling and antimicrobial properties. These wings are covered by periodic topography ranging from highly ordered hexagonal arrays of nanopillars to intricate “Christmas-tree” like structures with the ability to kill microbes by physically rupturing the cell membrane. In contrast, the topography of honeybee wings has received less attention. The role topography plays in antibiofouling, and antimicrobial activity of honeybee wings has never been investigated. Here, through antimicrobial and electron microscopy studies, we showed that pristine honeybee wings displayed no microbes on the wing surface. Also, the wings displayed antimicrobial properties that disrupt microbial cells and inhibit their growth. The antimicrobial activities of the wings were extremely effective at inhibiting the growth of Gram-negative bacterial cells when compared to Gram-positive bacterial cells. The fore wing was effective at inhibiting the growth of Gram-negative bacteria compared to Gram-positive samples. Electron microscopy revealed that the wings were studded with an array of rough, sharp, and pointed pillars that were distributed on both the dorsal and ventral sides, which enhanced anti-biofouling and antimicrobial effects. Our findings demonstrate the potential benefits of incorporating honeybee wings nanopatterns into the design of antibacterial nanomaterials which can be translated into countless applications in healthcare and industry.
Citation: Akamu J. Ewunkem, A'lyiha F. Beard, Brittany L. Justice, Sabrina L. Peoples, Jeffery A. Meixner, Watson Kemper, Uchenna B. Iloghalu. Honeybee wings hold antibiofouling and antimicrobial clues for improved applications in health care and industries[J]. AIMS Microbiology, 2023, 9(2): 332-345. doi: 10.3934/microbiol.2023018
Natural surfaces with remarkable properties and functionality have become the focus of intense research. Heretofore, the natural antimicrobial properties of insect wings have inspired research into their applications. The wings of cicadas, butterflies, dragonflies, and damselflies have evolved phenomenal anti-biofouling and antimicrobial properties. These wings are covered by periodic topography ranging from highly ordered hexagonal arrays of nanopillars to intricate “Christmas-tree” like structures with the ability to kill microbes by physically rupturing the cell membrane. In contrast, the topography of honeybee wings has received less attention. The role topography plays in antibiofouling, and antimicrobial activity of honeybee wings has never been investigated. Here, through antimicrobial and electron microscopy studies, we showed that pristine honeybee wings displayed no microbes on the wing surface. Also, the wings displayed antimicrobial properties that disrupt microbial cells and inhibit their growth. The antimicrobial activities of the wings were extremely effective at inhibiting the growth of Gram-negative bacterial cells when compared to Gram-positive bacterial cells. The fore wing was effective at inhibiting the growth of Gram-negative bacteria compared to Gram-positive samples. Electron microscopy revealed that the wings were studded with an array of rough, sharp, and pointed pillars that were distributed on both the dorsal and ventral sides, which enhanced anti-biofouling and antimicrobial effects. Our findings demonstrate the potential benefits of incorporating honeybee wings nanopatterns into the design of antibacterial nanomaterials which can be translated into countless applications in healthcare and industry.

| [1] |
Lesho EP, Laguio-Vila M (2019) The slow-motion catastrophe of antimicrobial resistance and practical interventions for all prescribers. Mayo Clin Proc 94: 1040-1047. https://doi.org/10.1016/j.mayocp.2018.11.005 
|
| [2] |
Murray CJL, Ikuta KS, Sharara F, et al. (2022) Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lance 399: 629-655. https://doi.org/10.1016/S0140-6736(21)02724-0
|
| [3] |
Saha M, Sarkar A (2021) Review on multiple facets of drug resistance: a rising challenge in the 21st century. J Xenobiot 11: 197-214. https://doi.org/10.3390/jox11040013
|
| [4] |
Gao W, Zhang L (2021) Nanomaterials arising amid antibiotic resistance. Nat Rev Microbiol 19: 5-6. https://doi.org/10.1038/s41579-020-00469-5
|
| [5] |
Oopath SV, Baji A, Abtahi M, et al. (2023) Nature-inspired biomimetic surfaces for controlling bacterial attachment and biofilm development. Adv Mater Interfaces 10: 2201425. https://doi.org/10.1002/admi.202201425
|
| [6] |
Wagner GP (2007) The developmental genetics of homology. Nat Rev Genet 8: 473-479.
|
| [7] | Linz DM, Tomoyasu Y (2018) Dual evolutionary origin of insect wings supported by an investigation of the abdominal wing serial homologs in Tribolium. Proc Natl Acad Sci 115: E658-E667. https://doi.org/10.1073/pnas.1711128115 |
| [8] |
Deora T, Sane SS, Sane SP (2021) Wings and halteres act as coupled dual oscillators in flies. Elife 10: e53824. https://doi.org/10.7554/eLife.53824
|
| [9] |
Salcedo MK, Socha JJ (2020) Circulation in insect wings. Integr Comp Biol 60: 1208-1220. https://doi.org/10.1093/icb/icaa124
|
| [10] |
Truong VK, Geeganagamage NM, Baulin VA, et al. (2017) The susceptibility of Staphylococcus aureus CIP 65.8 and Pseudomonas aeruginosa ATCC 9721 cells to the bactericidal action of nanostructured Calopteryx haemorrhoidalis damselfly wing surfaces. Appl Microbiol Biotechnol 101: 4683-4690. https://doi.org/10.1007/s00253-017-8205-9
|
| [11] |
Linklater DP, Juodkazis S, Rubanov S, et al. (2017) Comment on “bactericidal effects of natural nanotopography of dragonfly wing on Escherichia coli”. ACS Appl Mater Interfaces 9: 29387-29393. https://doi.org/10.1021/acsami.7b05707
|
| [12] |
Cheeseman S, Owen S, Truong VK, et al. (2018) Pillars of life: is there a relationship between lifestyle factors and the surface characteristics of dragonfly wings?. ACS Omega 3: 6039-6046. https://doi.org/10.1021/acsomega.8b00776
|
| [13] |
Hasan J, Webb HK, Truong VK, et al. (2013) Selective bactericidal activity of nanopatterned superhydrophobic cicada Psaltoda claripennis wing surfaces. Appl Microbiol Biotechnol 97: 9257-9262.
|
| [14] |
Nowlin K, Boseman A, Covell A, et al. (2015) Adhesion-dependent rupturing of Saccharomyces cerevisiae on biological antimicrobial nanostructured surfaces. J R Soc Interface 12. https://doi.org/10.1098/rsif.2014.0999
|
| [15] |
Ivanova EP, Hasan J, Webb HK, et al. (2012) Natural bactericidal surfaces: mechanical rupture of Pseudomonas aeruginosa cells by cicada wings. Small 8: 2489-2894. https://doi.org/10.1002/smll.201200528
|
| [16] |
Pogodin S, Hasan J, Baulin VA, et al. (2013) Biophysical model of bacterial cell interactions with nanopatterned cicada wing surfaces. Biophys J 104: 835-840. https://doi.org/10.1016/j.bpj.2012.12.046
|
| [17] |
Jenkins J, Mantell J, Neal C, et al. (2020) Antibacterial effects of nanopillar surfaces are mediated by cell impedance, penetration and induction of oxidative stress. Nat Commun 11: 1626. https://doi.org/10.1038/s41467-020-15471-x
|
| [18] |
Ivanova EP, Linklater DP, Werner M, et al. (2020) The multi-faceted mechano-bactericidal mechanism of nanostructured surfaces. Proc Natl Acad Sci 117: 12598-12605. https://doi.org/10.1073/pnas.1916680117
|
| [19] |
Heckmann TS, Schiffman JD (2019) Spatially organized nanopillar arrays dissimilarly affect the antifouling and antibacterial activities of Escherichia coli and Staphylococcus aureus. ACS Appl Nano Mater 3: 977-984. https://doi.org/10.1021/acsanm.9b01942
|
| [20] |
Bandara CD, Ballerin G, Leppanen M, et al. (2020) Resolving bio–nano interactions of E. coli bacteria–dragonfly wing interface with helium ion and 3D-structured illumination microscopy to understand bacterial death on nanotopography. ACS Biomater Sci Eng 6: 3925-3932. https://doi.org/10.1021/acsbiomaterials.9b01973
|
| [21] |
Dupont YL, Hansen DM, Olesen JM (2003) Structure of a plant-flower-visitor network in the high-altitude sub-alpine desert of Tenerife, Canary Islands. Ecography 26: 301-310. https://doi.org/10.1034/j.1600-0587.2003.03443.x
|
| [22] |
Greenleaf SS, Kremen C (2006) Wild bees enhance honey bees' pollination of hybrid sunflower. Proc Natl Acad Sci 103: 13890-13895. https://doi.org/10.1073/pnas.0600929103
|
| [23] |
Fratini F, Cilia G, Turchi B, et al. (2016) Beeswax: A minireview of its antimicrobial activity and its application in medicine. Asian Pac J Trop Med 9: 839-843. https://doi.org/10.1016/j.apjtm.2016.07.003
|
| [24] |
Rortais A, Barrucci F, Ercolano V, et al. (2021) A topic model approach to identify and track emerging risks from beeswax adulteration in the media. Food Control 119: 107435. https://doi.org/10.1016/j.foodcont.2020.107435
|
| [25] |
Nader RA, Mackieh R, Wehbe R, et al. (2021) Beehive products as antibacterial agents: a review. Antibiotics 10: 717. https://doi.org/10.3390/antibiotics10060717
|
| [26] |
Bang LM, Buntting C, Molan P (2003) The effect of dilution on the rate of hydrogen peroxide production in honey and its implications for wound healing. J Altern Complementary Med 9: 267-273. https://doi.org/10.1089/10755530360623383
|
| [27] |
Bizerra FC, Da Silva PI, Hayashi MAF (2012) Exploring the antibacterial properties of honey and its potential. Front Microbiol 3: 398. https://doi.org/10.3389/fmicb.2012.00398
|
| [28] |
Kuś PM, Szweda P, Jerković I, et al. (2016) Activity of Polish unifloral honeys against pathogenic bacteria and its correlation with colour, phenolic content, antioxidant capacity and other parameters. Lett Appl Microbiol 62: 269-276. https://doi.org/10.1111/lam.12541
|
| [29] |
Langlands Z, du Rand EE, Yusuf AA, et al. (2022) Functional response of the hypopharyngeal glands to a social parasitism challenge in Southern African honey bee subspecies. Parasitol Res 121: 267-274. https://doi.org/10.1007/s00436-021-07391-6
|
| [30] | Ma Y, Ning JG, Ren HL, et al. (2015) The function of resilin in honeybee wings. J Exp Biol 218: 2136-2142. https://doi.org/10.1242/jeb.117325 |
| [31] |
Ivanova EP, Hasan J, Webb HK, et al. (2013) Bactericidal activity of black silicon. Nat Commun 4. https://doi.org/2838. 10.1038/ncomms3838
|
| [32] |
Valiei A, Lin N, Bryche JF, et al. (2020) Mechano-bactericidal nanopillars require external forces to effectively kill bacteria. Nano Lett 20: 5720-5727. https://doi.org/10.1021/acs.nanolett.0c01343
|
| [33] |
Luo L, Zhou Y, Xu X, et al. (2020) Progress in construction of bio-inspired physico-antimicrobial surfaces. Nanotechnol Rev 9: 1562-1575. https://doi.org/10.1515/ntrev-2020-0089
|
| [34] |
Ivanova EP, Linklater DP, Aburto-Medina A, et al. (2021) Antifungal versus antibacterial defence of insect wings. J Colloid Interface Sci 603: 886-897. https://doi.org/10.1016/j.jcis.2021.06.093
|
| [35] |
Liang Y, Zhao J, Yan S (2017) Honeybees have hydrophobic wings that enable them to fly through fog and dew. J Bionic Eng 14: 549-556. https://doi.org/10.1016/S1672-6529(16)60415-8
|
| [36] |
Arias SL, Devorkin J, Spear JC, et al. (2020) Bacterial envelope damage inflicted by bioinspired nanostructures grown in a hydrogel. ACS Appl Bio Mate 3: 7974-7988. https://doi.org/10.1021/acsabm.0c01076
|
| [37] |
Hurtado-Rios JJ, Carrasco-Navarro U, Almanza-Pérez JC, et al. (2022) Ribosomes: the new role of ribosomal proteins as natural antimicrobials. Int J Mol Sci 23: 9123. https://doi.org/10.3390/ijms23169123
|
| [38] |
Yao X, Walter J, Burke S, et al. (2002) Atomic force microscopy and theoretical considerations of surface properties and turgor pressures of bacteria. Colloids Surf B 23: 213-230. https://doi.org/10.1016/S0927-7765(01)00249-1
|
| [39] |
Manina G, Dhar N, McKinney JD (2015) Stress and host immunity amplify Mycobacterium tuberculosis phenotypic heterogeneity and induce nongrowing metabolically active forms. Cell Host Microbe 17: 32-46. https://doi.org/10.1016/j.chom.2014.11.016
|
| [40] |
Priestman M, Thomas P, Robertson BD, et al. (2017) Mycobacteria modify their cell size control under sub-optimal carbon sources. Front Cell Dev Biol 5. https://doi.org/10.3389/fcell.2017.00064
|
| [41] |
Ewunkem AJ, Rodgers LS, Campbell D, et al. (2021) Experimental evolution of magnetite nanoparticle resistance in Escherichia coli. Nanomaterials 11. https://doi.org/10.3390/nano11030790
|
| [42] | Zhang X, Hao Y, Niu Q, et al. (2022) Division of labor among worker bees is associated with the lipidomic plasticity in their brains. Agriculture 12. https://doi.org/10.3390/agriculture12070952 |
| [43] |
da Luz GF, Santana WC, Santos CG, et al. (2022) Cuticle melanization and the expression of immune-related genes in the honeybee Apis mellifera (Hymenoptera: Apidae) adult workers. Comp Biochem Physiol Part B: Biochem Mol Biol 257: 110679. https://doi.org/10.1016/j.cbpb.2021.110679
|
| [44] |
Moore AJ, Breed MD, Moor MJ (1987) The guard honey bee: ontogeny and behavioural variability of workers performing a specialized task. Anim Behav 35: 1159-1167. https://doi.org/10.1016/S0003-3472(87)80172-0
|
| [45] |
Johnson BR (2005) Limited flexibility in the temporal caste system of the honey bee. Behav Ecol Sociobiol 58: 219-226. https://doi.org/10.1007/s00265-005-0949-z
|
| [46] |
Byhrø EMH, Salmela H, Vitlic A, et al. (2019) Different activation of immune-related genes in honey bee nurses and foragers (Apis mellifera). Apidologie 50: 463-471. https://doi.org/10.1007/s13592-019-00658-z
|
| [47] |
Vilmos P, Kurucz E (1998) Insect immunity: evolutionary roots of the mammalian innate immune system. Immunol Lett 62: 59-66. https://doi.org/10.1016/S0165-2478(98)00023-6
|
| [48] |
Kavanagh K, Reeves EP (2004) Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol Rev 28: 101-112. https://doi.org/10.1016/j.femsre.2003.09.002
|
| [49] |
Soares MPM, Elias-Neto M, Simões ZLP, et al. (2007) A cuticle protein gene in the honeybee: expression during development and in relation to the ecdysteroid titer. Insect Biochem Mol Biol 37: 1272-1282. https://doi.org/10.1016/j.ibmb.2007.07.014
|




Figures(5)

Akamu J. Ewunkem, A'lyiha F. Beard, Brittany L. Justice, Sabrina L. Peoples, Jeffery A. Meixner, Watson Kemper, Uchenna B. Iloghalu. Honeybee wings hold antibiofouling and antimicrobial clues for improved applications in health care and industries[J]. AIMS Microbiology, 2023, 9(2): 332-345. doi: 10.3934/microbiol.2023018







 DownLoad:
DownLoad: