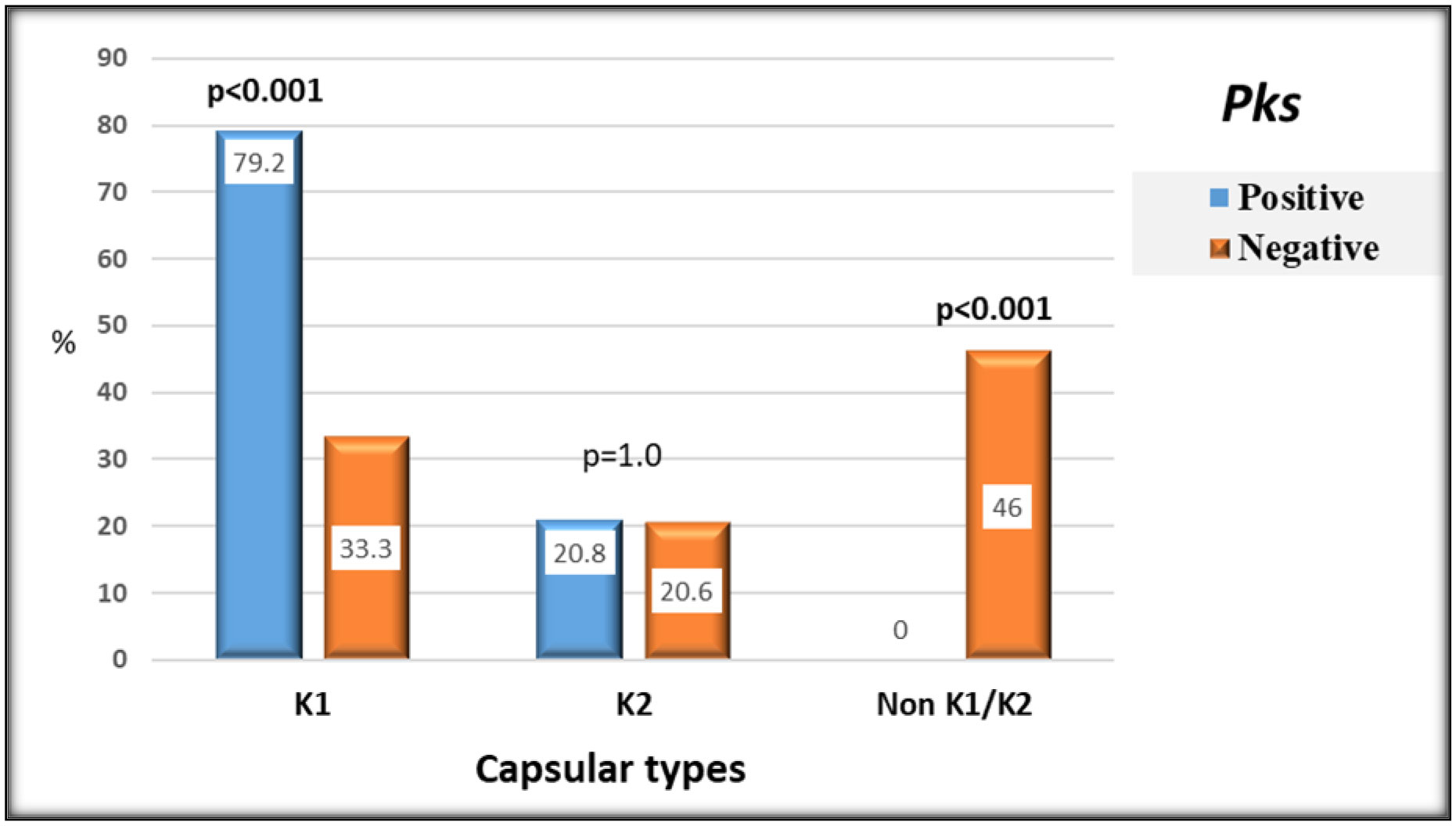
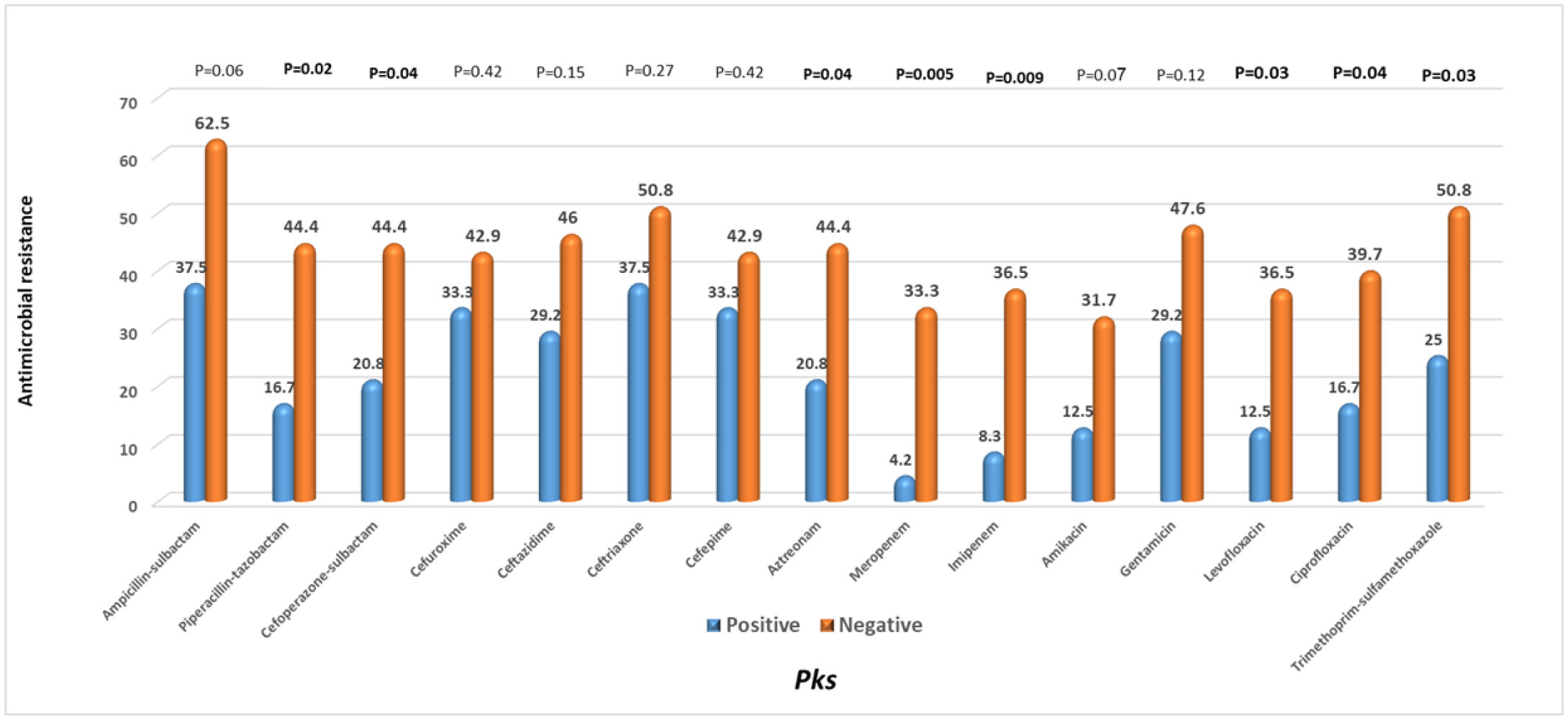
The pks genotoxic K. pneumoniae has recently triggered a widespread alarm. DNA damage and higher virulence have been linked to colibactin, a genotoxin expressed by the pks genomic island. Little is known about its molecular epidemiology in clinical isolates from Egypt. Therefore, this study was conducted to determine the prevalence and the microbiological and clinical features of pks harboring hospital-acquired K. pneumoniae isolates from Egypt. Eighty-seven hospital-acquired K. pneumoniae isolates from various specimen types were screened for pks colibactin island markers clbB, clbQ, clbA, and clbN by PCR. The pks-positive hvKp isolates were classified to one of the capsular types K1 and K2 using multiplex-PCR targeting K-serotype wzi and rmpA genes. The prevalence of pks+ strains was 27.6% (24/87). K1 capsular type, phenotypic, and genotypic hypervirulent isolates were significantly higher among pks+ strains than pks− strains (P < 0.001), while pks+ K. pneumoniae strains were found to be significantly less resistant to 8 of the antibiotic compounds tested than pks− strains. Carriage of K1 capsular type and mucoviscosity-associated rmp A gene and diabetes mellitus were identified to remain independent risk factors having a substantial association to pks-positivity by multivariate regression analysis. In conclusion, Hospital-acquired K. pneumoniae isolates in Egypt had an increased prevalence of the pks colibactin genotoxin. The significant occurrence of hypervirulent determinants in pks+ K. pneumoniae highlighted the genotoxin's possible pathogenicity combined with its distribution in several specimen types, which necessitates clinical attention and epidemic tracking.
Citation: Amira H. El-Ashry, Shimaa R. Hendawy, Noha Mostafa Mahmoud. Prevalence of pks genotoxin among hospital-acquired Klebsiella pneumoniae[J]. AIMS Microbiology, 2022, 8(1): 73-82. doi: 10.3934/microbiol.2022007
The pks genotoxic K. pneumoniae has recently triggered a widespread alarm. DNA damage and higher virulence have been linked to colibactin, a genotoxin expressed by the pks genomic island. Little is known about its molecular epidemiology in clinical isolates from Egypt. Therefore, this study was conducted to determine the prevalence and the microbiological and clinical features of pks harboring hospital-acquired K. pneumoniae isolates from Egypt. Eighty-seven hospital-acquired K. pneumoniae isolates from various specimen types were screened for pks colibactin island markers clbB, clbQ, clbA, and clbN by PCR. The pks-positive hvKp isolates were classified to one of the capsular types K1 and K2 using multiplex-PCR targeting K-serotype wzi and rmpA genes. The prevalence of pks+ strains was 27.6% (24/87). K1 capsular type, phenotypic, and genotypic hypervirulent isolates were significantly higher among pks+ strains than pks− strains (P < 0.001), while pks+ K. pneumoniae strains were found to be significantly less resistant to 8 of the antibiotic compounds tested than pks− strains. Carriage of K1 capsular type and mucoviscosity-associated rmp A gene and diabetes mellitus were identified to remain independent risk factors having a substantial association to pks-positivity by multivariate regression analysis. In conclusion, Hospital-acquired K. pneumoniae isolates in Egypt had an increased prevalence of the pks colibactin genotoxin. The significant occurrence of hypervirulent determinants in pks+ K. pneumoniae highlighted the genotoxin's possible pathogenicity combined with its distribution in several specimen types, which necessitates clinical attention and epidemic tracking.

| [1] |
Effah CY, Sun T, Liu S, et al. (2020) Klebsiella pneumoniae: an increasing threat to public health. Ann Clin Microbiol Antimicrob 19: 1. https://doi.10.1186/s12941-019-0343-8 
|
| [2] |
Lin ZW, Zheng JX, Bai B, et al. (2020) Characteristics of hypervirulent Klebsiella pneumoniae: Does low expression of rmpA contribute to the absence of Hypervirulence?. Front Microbiol 11: 436. https://doi.10.3389/fmicb.2020.00436
|
| [3] |
Zhu J, Wang T, Chen L, et al. (2021) Virulence factors in hypervirulent Klebsiella pneumoniae. Front Microbiol 12: 642484. https://doi.10.3389/fmicb.2021.642484
|
| [4] |
Choby JE, Howard-Anderson J, Weiss DS (2020) Hypervirulent Klebsiella pneumoniae-clinical and molecular perspectives. J Intern Med 287: 283-300. https://doi.10.1111/joim.13007
|
| [5] |
Nougayrède JP, Homburg S, Taieb F, et al. (2006) Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science 313: 848-51. https://doi.10.1126/science.1127059
|
| [6] |
McCarthy AJ, Martin P, Cloup E, et al. (2015) The genotoxin colibactin is a determinant of virulence in Escherichia coli K1 experimental neonatal systemic infection. Infect Immun 83: 3704-11. https://doi.10.1128/iai.00716-15
|
| [7] |
Shimpoh T, Hirata Y, Ihara S, et al. (2017) Prevalence of pks-positive Escherichia coli in Japanese patients with or without colorectal cancer. Gut Pathog 9: 35. https://doi.10.1186/s13099-017-0185-x
|
| [8] |
Putze J, Hennequin C, Nougayr'ede J P, et al. (2009) Genetic structure and distribution of the colibactin genomic island among members of the family Enterobacteriaceae. Infect. Immun 77: 4696-4703. https://doi.10.1128/IAI.00522-09
|
| [9] |
Struve C, Roe CC, Stegger M, et al. (2015) Mapping the evolution of Hypervirulent Klebsiella pneumoniae. mBio 6: e00630. https://doi.10.1128/mBio.00630-15
|
| [10] | Kreig N, Holt J (1984) Bergey's Manual of systemic bacteriology Vol.1. William and Wilkins, Baltimore, M.D. 21202 . USA: Williams & Wilkins Co 161-172. |
| [11] |
Lee HC, Chuang YC, Yu WL, et al. (2006) Clinical implications of hypermucoviscosity phenotype in Klebsiella pneumoniae isolates: association with invasive syndrome in patients with community-acquired bacteraemia. J Intern Med 259: 606-14. https://doi.10.1111/j.1365-2796.2006.01641.x
|
| [12] |
Ranjan A, Shaik S, Nandanwar N, et al. (2017) Comparative genomics of Escherichia coli isolated from skin and soft tissue and other Extraintestinal infections. mBio 8: e01070-17. https://doi.10.1128/mBio.01070-17
|
| [13] | Fialkina SV, Bondarenko VM, Naboka Iu L, et al. (2011) [Revealing the genetic determinants of Pks-pathogenicity island in clinical strains of enterobacteria]. Zh Mikrobiol Epidemiol Immunobiol 1: 3-7. Available from: https://pubmed.ncbi.nlm.nih.gov/22145340/ |
| [14] |
Compain F, Babosan A, Brisse S, et al. (2014) Multiplex PCR for detection of seven virulence factors and K1/K2 capsular serotypes of Klebsiella pneumoniae. J Clin Microbiol 52: 4377-80. https://doi.10.1128/JCM.02316-14
|
| [15] | Patel JB, Cockerill F, Bradford PA (2015) Performance standards for antimicrobial susceptibility testing: twenty-fifth informational supplement. CLSI Document M100-S25 . |
| [16] |
Magiorakos AP, Srinivasan A, Carey RB, et al. (2012) Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 18: 268-81. https://doi.10.1111/j.1469-0691.2011.03570.x
|
| [17] |
Cuevas-Ramos G, Petit CR, Marcq I, et al. (2010) Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc Natl Acad Sci USA 107: 11537-42. https://doi.10.1073/pnas.1001261107
|
| [18] |
Lan Y, Zhou M, Jian Z, et al. (2019) Prevalence of pks gene cluster and characteristics of Klebsiella pneumoniae-induced bloodstream infections. J Clin Lab Anal 33: e22838. https://doi.10.1002/jcla.22838
|
| [19] |
Lai YC, Lin AC, Chiang MK, et al. (2014) Genotoxic Klebsiella pneumoniae in Taiwan. PLoS One 9: e96292. https://doi.10.1371/journal.pone.0096292
|
| [20] |
Johnson JR, Johnston B, Kuskowski MA, et al. (2008) Molecular epidemiology and phylogenetic distribution of the Escherichia coli pks genomic island. J Clin Microbiol 46: 3906-11. https://doi.10.1128/jcm.00949-08
|
| [21] |
Yeh KM, Kurup A, Siu LK, et al. (2007) Capsular serotype K1 or K2, rather than magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in Singapore and Taiwan. J Clin Microbiol 45: 466-71. https://doi.10.1128/jcm.01150-06
|
| [22] |
Strakova N, Korena K, Karpiskova R (2021) Klebsiella pneumoniae producing bacterial toxin colibactin as a risk of colorectal cancer development-A systematic review. Toxicon 197: 126-135. https://doi.10.1016/j.toxicon.2021.04.007
|
| [23] |
Xu Q, Yang X, Chan EWC, et al. (2021) The hypermucoviscosity of hypervirulent K. pneumoniae confers the ability to evade neutrophil-mediated phagocytosis. Virulence 12: 2050-2059. https://doi.10.1080/21505594.2021.1960101
|
| [24] |
Yu WL, Ko WC, Cheng KC, et al. (2008) Comparison of prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with capsular K1/K2 and non-K1/K2 serotypes. Diagn Microbiol Infect Dis 62: 1-6. https://doi.10.1016/j.diagmicrobio.2008.04.007
|
| [25] |
Yu WL, Ko WC, Cheng KC, et al. (2006) Association between rmpA and magA genes and clinical syndromes caused by Klebsiella pneumoniae in Taiwan. Clin Infect Dis 42: 1351-8. https://doi.10.1086/503420
|
| [26] |
Shi Q, Quan J, Lan P, et al. (2020) Prevalence and characteristics of pks gene cluster harbouring Klebsiella pneumoniae from bloodstream infection in China. Epidemiol Infect 148: e69. https://doi.10.1017/s0950268820000655
|
| [27] |
Siu LK, Yeh KM, Lin JC, et al. (2012) Klebsiella pneumoniae liver abscess: a new invasive syndrome. Lancet Infect Dis 12: 881-7. https://doi.10.1016/s1473-3099(12)70205-0
|
| [28] |
Fang CT, Lai SY, Yi WC, et al. (2007) Klebsiella pneumoniae genotype K1: an emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin Infect Dis 45: 284-93. https://doi.10.1086/519262
|
| [29] |
Bialek-Davenet S, Criscuolo A, Ailloud F, et al. (2014) Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg Infect Dis 20: 1812-20. https://doi.10.3201/eid2011.140206
|
| [30] |
Li W, Sun G, Yu Y, et al. (2014) Increasing occurrence of antimicrobial-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in China. Clin Infect Dis 58: 225-32. https://doi.10.1093/cid/cit675
|
Figures(2) / Tables(3)

Amira H. El-Ashry, Shimaa R. Hendawy, Noha Mostafa Mahmoud. Prevalence of pks genotoxin among hospital-acquired Klebsiella pneumoniae[J]. AIMS Microbiology, 2022, 8(1): 73-82. doi: 10.3934/microbiol.2022007







 DownLoad:
DownLoad: