Citation: Ashrafus Safa, Jinath Sultana Jime, Farishta Shahel. Cholera toxin phage: structural and functional diversity between Vibrio cholerae biotypes[J]. AIMS Microbiology, 2020, 6(2): 144-151. doi: 10.3934/microbiol.2020009

| [1] |
Kaper JB, Morris JG, Levine MM (1995) Cholera. Clin Microbiol Rev 8: 48-86. doi: 10.1128/CMR.8.1.48

|
| [2] |
Albert M, Ansaruzzaman M, Bardhan P, et al. (1993) Large epidemic of cholera-like disease in Bangladesh caused by Vibrio cholerae 0139 synonym Bengal. The Lancet 342: 387-390. doi: 10.1016/0140-6736(93)91906-3
|
| [3] |
Sánchez J, Holmgren J (2008) Cholera toxin structure, gene regulation and pathophysiological and immunological aspects. Cell Mol Life Sci 65: 1347-1360. doi: 10.1007/s00018-008-7496-5
|
| [4] |
Olsvik Ø, Wahlberg J, Petterson B, et al. (1993) Use of automated sequencing of polymerase chain reaction-generated amplicons to identify three types of cholera toxin subunit B in Vibrio cholerae O1 strains. J Clin Microbiol 31: 22-25. doi: 10.1128/JCM.31.1.22-25.1993
|
| [5] | Mekalanos JJ (1985) Cholera toxin: genetic analysis, regulation and role in pathogenesis. Curr Top Microbiol Immunol 118: 97-118. |
| [6] |
Waldor MK, Mekalanos JJ (1996) Lysogenic conversion by a filamentous bacteriophage encoding cholera toxin. Science 272: 1910-1914. doi: 10.1126/science.272.5270.1910
|
| [7] |
Waldor MK, Rubin EJ, Pearson GDN, et al. (1997) Regulation, replication, and integration functions of the Vibrio cholerae CTXφ are encoded by regions RS2. Mol Microbiol 24: 917-926. doi: 10.1046/j.1365-2958.1997.3911758.x
|
| [8] |
Huber KE, Waldor MK (2002) Filamentous phage integration requires the host recombinases XerC and XerD. Nature 417: 656-659. doi: 10.1038/nature00782
|
| [9] |
Davis BM, Waldor MK (2003) Filamentous phages linked to virulence of Vibrio cholerae. Curr Opin Microbiol 6: 35-42. doi: 10.1016/S1369-5274(02)00005-X
|
| [10] |
Trucksis M, Galen JE, Michalski J, et al. (1993) Accessory cholera enterotoxin (Ace), the third toxin of a Vibrio cholerae virulence cassette. Proc Natl Acad Sci USA 90: 5267-5271. doi: 10.1073/pnas.90.11.5267
|
| [11] |
Faruque SM, Albert MJ, Mekalanos JJ, et al. (1998) Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae. Microbiol Mol Biol Rev 62: 1301-1314. doi: 10.1128/MMBR.62.4.1301-1314.1998
|
| [12] |
Heilpern AJ, Waldor MK (2003) pIIICTX, a predicted CTXφ minor coat protein, can expand the host range of coliphage fd to include Vibrio cholerae. J Bacteriol 185: 1037-1044. doi: 10.1128/JB.185.3.1037-1044.2003
|
| [13] |
Kimsey HH, Nair GB, Ghosh A, et al. (1998) Diverse CTXφ and evolution of new pathogenic Vibrio cholerae. Lancet 352: 457-458. doi: 10.1016/S0140-6736(05)79193-5
|
| [14] |
Davis BM, Kimsey HH, Kimsey WC, et al. (1999) The Vibrio cholerae O139 Calcutta bacteriophage CTXφ is infectious and encodes a novel repressor. J Bacteriol 181: 6779-6787. doi: 10.1128/JB.181.21.6779-6787.1999
|
| [15] |
Mukhopadhyay AK, Chakaraborty S, Takeda Y, et al. (2001) Characterization of VPI pathogenicity island and CTXφ prophage in environmental strains of Vibrio cholerae. J Bacteriol 183: 4737-474. doi: 10.1128/JB.183.16.4737-4746.2001
|
| [16] |
Bhattacharya T, Chatterjee S, Maiti D, et al. (2006) Molecular analysis of the rstR and orfU genes of the CTX prophages integrated in the small chromosomes of environmental Vibrio cholerae non-O1, non-O139 strains. Environ Microbiol 8: 526-534. doi: 10.1111/j.1462-2920.2005.00932.x
|
| [17] |
Nusrin S, Khan GY, Bhuiyan NA, et al. (2004) Diverse CTX phages among toxigenic Vibrio cholerae O1 and O139 strains isolated between 1994 and 2002 in an area where cholera is endemic in Bangladesh. J Clin Microbiol 42: 5854-5856. doi: 10.1128/JCM.42.12.5854-5856.2004
|
| [18] |
Kimsey H, Waldor MK (1998) CTXφ immunity: application in the development of cholera vaccines. Proc Natl Acad Sci USA 95: 7035-7039. doi: 10.1073/pnas.95.12.7035
|
| [19] |
Davis MB, Moyer KE, Boyd EF, et al. (2000) CTX prophages in classical biotype Vibrio cholerae: functional phage genes but dysfunctional phage genomes. J Bacteriol 182: 6992-6998. doi: 10.1128/JB.182.24.6992-6998.2000
|
| [20] |
Faruque SM, Tam VC, Chowdhury N, et al. (2007) Genomic analysis of the Mozambique strain of Vibrio cholerae O1 reveals the origin of El Tor strains carrying classical CTX prophage. Proc Natl Acad Sci USA 104: 5151-5156. doi: 10.1073/pnas.0700365104
|
| [21] |
Mekalanos JJ (1983) Duplication and amplification of toxin genes in Vibrio cholerae. Cell 35: 253-263. doi: 10.1016/0092-8674(83)90228-3
|
| [22] |
Trucksis M, Michalsky J, Deng YK, et al. (1998) The Vibrio cholerae genome contains two unique circular chromosomes. Proc Natl Acad Sci USA 95: 14464-14469. doi: 10.1073/pnas.95.24.14464
|
| [23] |
Nandi S, Maiti D, Saha A, et al. (2003) Genesis of variants of Vibrio cholerae O1 biotype El Tor: role of the CTXφ array and its position in the genome. Microbiology 149: 89-97. doi: 10.1099/mic.0.25599-0
|
| [24] |
Lee JH, Han KH, Choi SY, et al. (2006) Multilocus sequence typing (MLST) analysis of Vibrio cholerae O1 El Tor isolates from Mozambique that harbour the classical CTX prophage. J Med Microbiol 55: 165-170. doi: 10.1099/jmm.0.46287-0
|
| [25] |
Faruque SM, Asadulghani, Kamruzzaman M, et al. (2002) RS1 element of Vibrio cholerae can propagate horizontally as a filamentous phage exploiting the morphogenesis genes of CTXphi. Infect Immun 70: 163-170. doi: 10.1128/IAI.70.1.163-170.2002
|
| [26] |
Faruque SM, Kamruzzaman M, Asadulghan, et al. (2003) CTXφ-independent production of the RS1 satellite phage by Vibrio cholerae. Proc Natl Acad Sci USA 100: 1280-1285. doi: 10.1073/pnas.0237385100
|
| [27] |
Davis BM, Kimsey HH, Kane AV, et al. (2002) A satellite phage-encoded antirepressor induces repressor aggregation and cholera toxin gene transfer. EMBO J 21: 4240-4249. doi: 10.1093/emboj/cdf427
|
| [28] |
Safa A, Nair GB, Kong RYC (2010) Evolution of new variants of Vibrio cholerae O1. Trends Microbiol 18: 46-54. doi: 10.1016/j.tim.2009.10.003
|
| [29] |
Finkelstein RA, Burks F, Zupan A, et al. (1987) Epitopes of the cholera family of enterotoxins. Rev Infect Dis 9: 544-561. doi: 10.1093/clinids/9.3.544
|
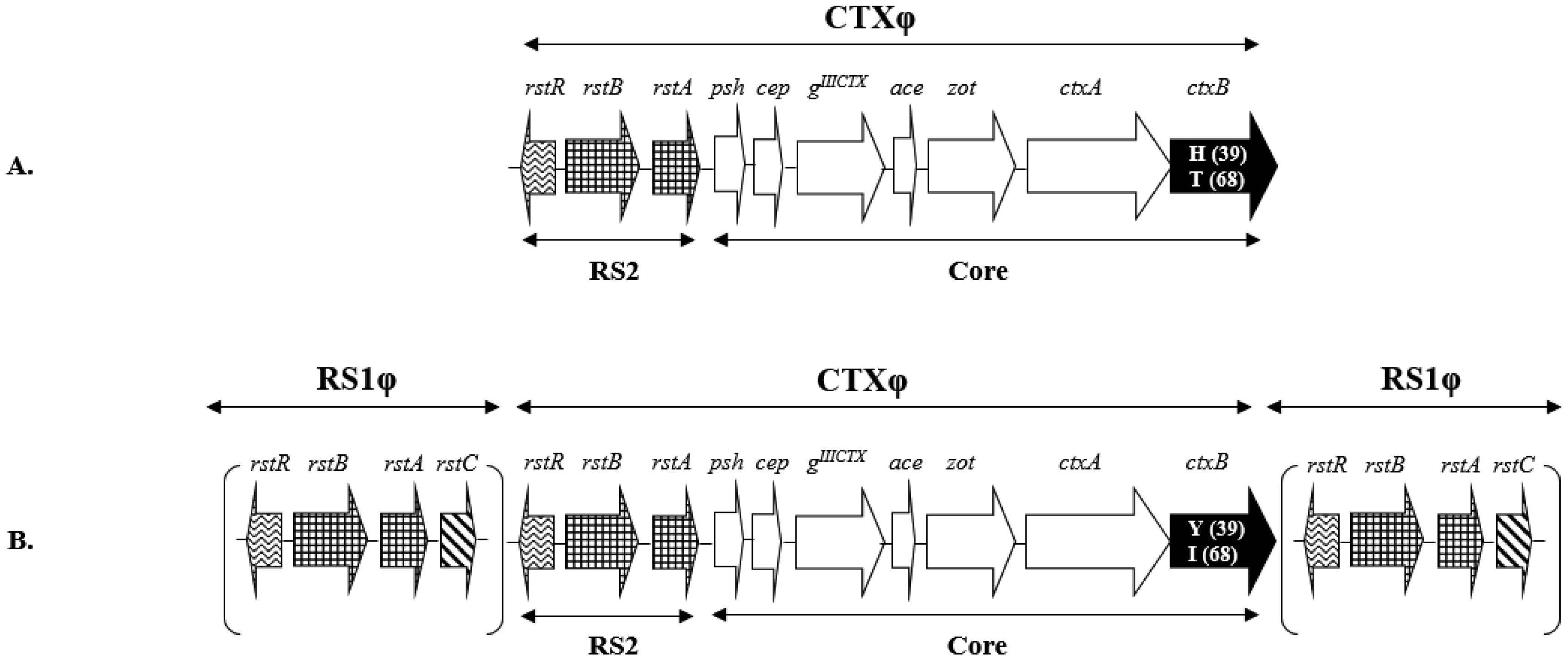

Figures(2)

Ashrafus Safa, Jinath Sultana Jime, Farishta Shahel. Cholera toxin phage: structural and functional diversity between Vibrio cholerae biotypes[J]. AIMS Microbiology, 2020, 6(2): 144-151. doi: 10.3934/microbiol.2020009







 DownLoad:
DownLoad: