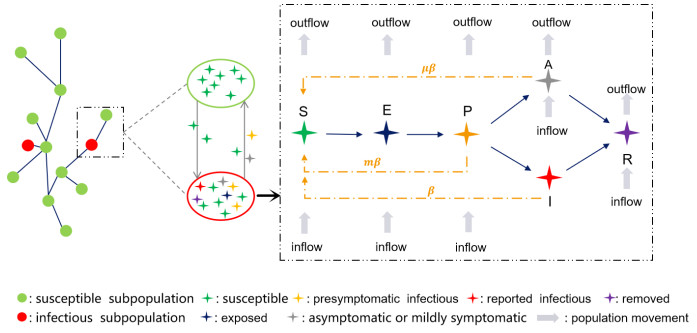
The emergence of many presymptomatic hidden transmission events significantly complicated the intervention and control of the spread of COVID-19 in the USA during the year 2020. To analyze the role that presymptomatic infections play in the spread of this disease, we developed a state-level metapopulation model to simulate COVID-19 transmission in the USA in 2020 during which period the number of confirmed cases was more than in any other country. We estimated that the transmission rate (i.e., the number of new infections per unit time generated by an infected individual) of presymptomatic infections was approximately 59.9% the transmission rate of reported infections. We further estimated that {at any point in time the} average proportion of infected individuals in the presymptomatic stage was consistently over 50% of all infected individuals. Presymptomatic transmission was consistently contributing over 52% to daily new infections, as well as consistently contributing over 50% to the effective reproduction number from February to December. Finally, non-pharmaceutical intervention targeting presymptomatic infections was very effective in reducing the number of reported cases. These results reveal the significant contribution that presymptomatic transmission made to COVID-19 transmission in the USA during 2020, as well as pave the way for the design of effective disease control and mitigation strategies.
Citation: Luyu Zhang, Zhaohua Zhang, Sen Pei, Qing Gao, Wei Chen. Quantifying the presymptomatic transmission of COVID-19 in the USA[J]. Mathematical Biosciences and Engineering, 2024, 21(1): 861-883. doi: 10.3934/mbe.2024036
The emergence of many presymptomatic hidden transmission events significantly complicated the intervention and control of the spread of COVID-19 in the USA during the year 2020. To analyze the role that presymptomatic infections play in the spread of this disease, we developed a state-level metapopulation model to simulate COVID-19 transmission in the USA in 2020 during which period the number of confirmed cases was more than in any other country. We estimated that the transmission rate (i.e., the number of new infections per unit time generated by an infected individual) of presymptomatic infections was approximately 59.9% the transmission rate of reported infections. We further estimated that {at any point in time the} average proportion of infected individuals in the presymptomatic stage was consistently over 50% of all infected individuals. Presymptomatic transmission was consistently contributing over 52% to daily new infections, as well as consistently contributing over 50% to the effective reproduction number from February to December. Finally, non-pharmaceutical intervention targeting presymptomatic infections was very effective in reducing the number of reported cases. These results reveal the significant contribution that presymptomatic transmission made to COVID-19 transmission in the USA during 2020, as well as pave the way for the design of effective disease control and mitigation strategies.

| [1] | World Health Organization, Coronavirus disease 2019 (COVID-19): Situation report, 2020. Available from: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports |
| [2] | World Health Organization, WHO Coronavirus (COVID-19) Dashboard, 2021. Available from: https://covid19.who.int 2021 |
| [3] |
M. L. Holshue, C. DeBolt, S. Lindquist, K. H. Lofy, J. Wiesman, H. Bruce, et al., First case of 2019 novel coronavirus in the United States, N. Engl. J. Med., 382 2020,929–936. https://doi.org/10.1056/NEJMoa2001191 doi: 10.1056/NEJMoa2001191

|
| [4] |
E. Dong, H. Du, L. Gardner, An interactive web-based dashboard to track COVID-19 in real time, Lancet Infect. Dis., 20 (2020), 533–534. https://doi.org/10.1016/S1473-3099(20)30120-1 doi: 10.1016/S1473-3099(20)30120-1
|
| [5] |
A. L. Rasmussen, S. V. Popescu, SARS-CoV-2 transmission without symptoms, Science, 371 (2021), 1206–1207. https://doi.org/10.1126/science.abf9569 doi: 10.1126/science.abf9569
|
| [6] |
L. Tian, X. Li, F. Qi, Q. Y. Tang, V. Tang, J. Liu, et al., Harnessing peak transmission around symptom onset for non-pharmaceutical intervention and containment of the COVID-19 pandemic, Nat. Commun., 12 (2021), 1147. https://doi.org/10.1038/s41467-021-21385-z doi: 10.1038/s41467-021-21385-z
|
| [7] |
D. P. Oran, E. J. Topol, Prevalence of asymptomatic SARS-CoV-2 infection, Ann. Int. Med., 174 (2021), 286–287. https://doi.org/10.7326/L20-1285 doi: 10.7326/L20-1285
|
| [8] |
S. W. Park, D. M. Cornforth, J. Dushoff, J. S. Weitz, The time scale of asymptomatic transmission affects estimates of epidemic potential in the COVID-19 outbreak, Epidemics, 31 (2020), 100392. https://doi.org/10.1016/j.epidem.2020.100392 doi: 10.1016/j.epidem.2020.100392
|
| [9] | K. Mizumoto, K. Kagaya, A. Zarebski, G. Chowell, Estimating the asymptomatic proportion of coronavirus disease 2019 (COVID-19) cases on board the Diamond Princess cruise ship, Yokohama, Japan, 2020, Eurosurveillance, 25 (2020), 2000180. |
| [10] |
J. F. W. Chan, S. Yuan, K. H. Kok, K. K. W. To, H. Chu, J. Yang, et al., A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster, Lancet, 395 (2020), 514–523. https://doi.org/10.1016/S0140-6736(20)30154-9 doi: 10.1016/S0140-6736(20)30154-9
|
| [11] |
L. Ferretti, C. Wymant, M. Kendall, L. Zhao, A. Nurtay, L. Abeler-Dörner, et al., Quantifying SARS-CoV-2 transmission suggests epidemic control with digital contact tracing, Science, 368 (2020), eabb6936. https://doi.org/10.1126/science.abb6936 doi: 10.1126/science.abb6936
|
| [12] | Z. D. Tong, A. Tang, K. F. Li, P. Li, H. L. Wang, J. P. Yi, et al., Potential presymptomatic transmission of SARS-CoV-2, Zhejiang province, China, 2020, Emerg. Infect. Dis., 26 (2020), 1052. https://doi.org/10.3201%2Feid2605.200198 |
| [13] |
S. J. Krieg, J. J. Schnur, M. L. Miranda, M. E. Pfrender, N. V. Chawla, Symptomatic, presymptomatic, and asymptomatic transmission of SARS-CoV-2 in a university student population, August—November 2020, Public Health Rep., 137 (2022), 1023–1030. https://doi.org/10.1177/00333549221110300 doi: 10.1177/00333549221110300
|
| [14] |
X. He, E. H. Y. Lau, P. Wu, X. Deng, J. Wang, X. Hao, et al., Temporal dynamics in viral shedding and transmissibility of COVID-19, Nat. Med., 26 (2020), 672–675. https://doi.org/10.1038/s41591-020-0869-5 doi: 10.1038/s41591-020-0869-5
|
| [15] |
D. Buitrago-Garcia, D. Egli-Gany, M. J. Counotte, S. Hossmann, H. Imeri, A. M. Ipekci, et al., Occurrence and transmission potential of asymptomatic and presymptomatic SARS-CoV-2 infections: A living systematic review and meta-analysis, PLoS Med., 17 (2020), e1003346. https://doi.org/10.1371/journal.pmed.1003346 doi: 10.1371/journal.pmed.1003346
|
| [16] |
S. M. Moghadas, M. C. Fitzpatrick, P. Sah, A. Pandey, A. Shoukat, B. H. Singer, et al., The implications of silent transmission for the control of COVID-19 outbreaks, Proc. Natl. Acad. Sci. USA, 117 (2020, 17513–17515. https://doi.org/10.1073/pnas.2008373117 doi: 10.1073/pnas.2008373117
|
| [17] |
L. C. Tindale, J. E. Stockdale, M. Coombe, E. S. Garlock, W. Y. V. Lau, M. Saraswat, et al., Evidence for transmission of COVID-19 prior to symptom onset, Elife, 9 (2020), e57149. https://doi.org/10.7554/eLife.57149 doi: 10.7554/eLife.57149
|
| [18] |
Z. Du, X. Xu, Y. Wu, L. Wang, B. J. Cowling, L. A. Meyers, Serial interval of COVID-19 among publicly reported confirmed cases, Emerg. Infect. Dis., 26 (2020), 1341. https://doi.org/10.3201/eid2606.200357 doi: 10.3201/eid2606.200357
|
| [19] |
S. A. McDonald, F. Miura, E. R. A. Vos, M. van Boven, H. E. de Melker, F. R. M. van der Klis, et al., Estimating the asymptomatic proportion of SARS-CoV-2 infection in the general population: Analysis of nationwide serosurvey data in the Netherlands, Eur. J. Epidemiol., 36 (2021), 735–739. https://doi.org/10.1007/s10654-021-00768-y doi: 10.1007/s10654-021-00768-y
|
| [20] |
W. E. Wei, Z. Li, C. J. Chiew, S. E. Yong, M. P. Toh, V. J. Lee, Presymptomatic transmission of SARS-CoV-2—Singapore, january 23—march 16, 2020, Morb. Mortal. Wkly. Rep., 69 (2020), 411. https://doi.org/10.3724/SP.J.1123.2021.03024 doi: 10.3724/SP.J.1123.2021.03024
|
| [21] |
C. Martínez, H. Serrano-Coll, Á. Faccini, V. Contreras, K. Galeano, Y. Botero, et al., SARS-CoV-2 in a tropical area of Colombia, a remarkable conversion of presymptomatic to symptomatic people impacts public health, BMC Infect. Dis., 22 (2022), 1–5. https://doi.org/10.1186/s12879-022-07575-0 doi: 10.1186/s12879-022-07575-0
|
| [22] |
R. Subramanian, Q. He, M. Pascual, Quantifying asymptomatic infection and transmission of COVID-19 in New York City using observed cases, serology, and testing capacity, Proc. Natl. Acad. Sci. USA, 118 (2021), e2019716118. https://doi.org/10.1073/pnas.2019716118 doi: 10.1073/pnas.2019716118
|
| [23] |
X. Hao, S. Cheng, D. Wu, T. Wu, X. Lin, C. Wang, Reconstruction of the full transmission dynamics of COVID-19 in Wuhan, Nature, 584 (2020), 420–424. https://doi.org/10.1038/s41586-020-2554-8 doi: 10.1038/s41586-020-2554-8
|
| [24] |
R. Li, S. Pei, B. Chen, Y. Song, T. Zhang, W. Yang, et al., Substantial undocumented infection facilitates the rapid dissemination of novel coronavirus (SARS-CoV-2), Science, 368 (2020), 489–493. https://doi.org/10.1126/science.abb3221 doi: 10.1126/science.abb3221
|
| [25] |
M. Monod, A. Blenkinsop, X. Xi, D. Hebert, S. Bershan, S. Tietze, et al., Age groups that sustain resurging COVID-19 epidemics in the United States, Science, 371 (2021), eabe8372. https://doi.org/10.1126/science.abe8372 doi: 10.1126/science.abe8372
|
| [26] |
S. Pei, S. Kandula, W. Yang, J. Shaman, Forecasting the spatial transmission of influenza in the United States, Proc. Natl. Acad. Sci. USA, 115 (2018), 2752–2757. https://doi.org/10.1073/pnas.1708856115 doi: 10.1073/pnas.1708856115
|
| [27] |
S. Pei, S. Kandula, J. Shaman, Differential effects of intervention timing on COVID-19 spread in the United States, Sci. Adv., 6 (2020), eabd6370. https://doi.org/10.1126/sciadv.abd6370 doi: 10.1126/sciadv.abd6370
|
| [28] |
J. L. Anderson, An ensemble adjustment kalman filter for data assimilation, Mon, Weather Rev., 129 (2001), 2884–2903. https://doi.org/10.1175/1520-0493(2001)129<2884:AEAKFF>2.0.CO;2 doi: 10.1175/1520-0493(2001)129<2884:AEAKFF>2.0.CO;2
|
| [29] |
J. Shaman, A. Karspeck, Forecasting seasonal outbreaks of influenza, Proc. Natl. Acad. Sci. USA, 109 (2012), 20425–20430. https://doi.org/10.1073/pnas.1208772109 doi: 10.1073/pnas.1208772109
|
| [30] | S. Pei, T. K. Yamana, S. Kandula, M. Galanti, J. Shaman, Overall burden and characteristics of COVID-19 in the United States during 2020, preprint, medRxiv: 2021.02. https://doi.org/10.1101/2021.02.15.21251777 |
| [31] |
M. S. Arulampalam, S. Maskell, N. Gordon, T. Clapp, A tutorial on particle filters for online nonlinear/non-Gaussian Bayesian tracking, IEEE Trans. Signal Process., 50 (2002), 174–188. https://doi.org/10.1109/78.978374 doi: 10.1109/78.978374
|
| [32] |
C. Snyder, T. Bengtsson, P. Bickel, J. Anderson, Obstacles to high-dimensional particle filtering, Mon. Weather Rev., 136 (2008), 4629–4640. https://doi.org/10.1175/2008MWR2529.1 doi: 10.1175/2008MWR2529.1
|
| [33] |
W. Yang, M. Lipsitch, J. Shaman, Inference of seasonal and pandemic influenza transmission dynamics, Proc. Natl. Acad. Sci. USA, 112 (2015), 2723–2728. https://doi.org/10.1073/pnas.1415012112 doi: 10.1073/pnas.1415012112
|
| [34] |
O. Diekmann, J. Heesterbeek, M. G. Roberts, The construction of next-generation matrices for compartmental epidemic models, J. R. Soc. Interface, 7 (2010), 873–885. https://doi.org/10.1098/rsif.2009.0386 doi: 10.1098/rsif.2009.0386
|
| [35] |
J. L. Daly, B. Simonetti, K. Klein, K. E. Chen, M. K. Williamson, C. Antón-Plágaro, et al., Neuropilin-1 is a host factor for SARS-CoV-2 infection, Science, 370 (2020), 861–865. https://doi.org/10.1126/science.abd3072 doi: 10.1126/science.abd3072
|
| [36] | S. Sanche, Y. T. Lin, C. Xu, E. Romero-Severson, N. Hengartner, R. Ke, High contagiousness and rapid spread of severe acute respiratory syndrome coronavirus 2, Emerg. Inf. Dis., 26 (2020), 1470. https://doi.org/10.3201%2Feid2607.200282 |
| [37] |
S. Lai, N. W. Ruktanonchai, L. Zhou, O. Prosper, W. Luo, J. R. Floyd, et al., Effect of non-pharmaceutical interventions to contain COVID-19 in China, Nature, 585 (2020), 410–413. https://doi.org/10.1038/s41586-020-2293-x doi: 10.1038/s41586-020-2293-x
|
| [38] |
M. Chinazzi, J. T. Davis, M. Ajelli, C. Gioannini, M. Litvinova, S. Merler, et al., The effect of travel restrictions on the spread of the 2019 novel coronavirus (COVID-19) outbreak, Science, 368 (2020), 395–400. https://doi.org/10.1126/science.aba9757 doi: 10.1126/science.aba9757
|
| [39] |
H. Tian, Y. Liu, Y. Li, C. H. Wu, B. Chen, M. U. Kraemer, et al., An investigation of transmission control measures during the first 50 days of the COVID-19 epidemic in China, Science, 368 (2020), 638–642. https://doi.org/10.1126/science.abb6105 doi: 10.1126/science.abb6105
|
| [40] |
B. F. Maier, D. Brockmann, Effective containment explains subexponential growth in recent confirmed COVID-19 cases in China, Science, 368 (2020), 742–746. https://doi.org/10.1126/science.abb4557 doi: 10.1126/science.abb4557
|
| [41] |
J. Zhang, M. Litvinova, Y. Liang, Y. Wang, W. Wang, S. Zhao, et al., Changes in contact patterns shape the dynamics of the COVID-19 outbreak in China, Science, 368 (2020), 1481–1486. https://doi.org/10.1126/science.abb8001 doi: 10.1126/science.abb8001
|
| [42] |
M. U. Kraemer, C. H. Yang, B. Gutierrez, C. H. Wu, B. Klein, D. M. Pigott, et al., The effect of human mobility and control measures on the COVID-19 epidemic in China, Science, 368, (2020), 493–497. https://doi.org/10.1126/science.abb4218 doi: 10.1126/science.abb4218
|
| [43] |
J. Dehning, J. Zierenberg, F. P. Spitzner, M. Wibral, J. P. Neto, M. Wilczek, V. Priesemann, Inferring change points in the spread of COVID-19 reveals the effectiveness of interventions, Science, 369 (2020), eabb9789. https://doi.org/10.1126/science.abb9789 doi: 10.1126/science.abb9789
|
| [44] |
M. Du, Contact tracing as a measure to combat COVID-19 and other infectious diseases, Am. J. Infect. Control, 50 (2022), 638–644. https://doi.org/10.1016/j.ajic.2021.11.031 doi: 10.1016/j.ajic.2021.11.031
|
| [45] |
S. Liu, T. Yamamoto, Role of stay-at-home requests and travel restrictions in preventing the spread of COVID-19 in Japan, Transp. Res. Part A, 159 (2022), 1–16. https://doi.org/10.1016/j.tra.2022.03.009 doi: 10.1016/j.tra.2022.03.009
|
| [46] |
N. G. Davies, P. Klepac, Y. Liu, K. Prem, M. Jit, R. M. Eggo, Age-dependent effects in the transmission and control of COVID-19 epidemics, Nat. Med., 26 (2020), 1205–1211. https://doi.org/10.1038/s41591-020-0962-9 doi: 10.1038/s41591-020-0962-9
|
| [47] |
A. James, M. J. Plank, R. N. Binny, A. Lustig, K. Hannah, S. Hendy, N. Steyn, A structured model for COVID-19 spread: Modelling age and healthcare inequities, Math. Med. Biol., 38 (2021), 299–313. https://doi.org/10.1093/imammb/dqab006 doi: 10.1093/imammb/dqab006
|
| [48] |
E. A. Undurraga, G. Chowell, K. Mizumoto, COVID-19 case fatality risk by age and gender in a high testing setting in Latin America: Chile, March–August 2020, Infect. Dis. Poverty, 10 (2021), 1–11. https://doi.org/10.1186/s40249-020-00785-1 doi: 10.1186/s40249-020-00785-1
|
| [49] |
R. Verity, L. Okell, I. Dorigatti, P. Winskill, C. Whittaker, N. Imai, et al., Estimates of the severity of coronavirus disease 2019: A model-based analysis, Lancet Infect. Dis., 20 (2020), 669–677. https://doi.org/10.1016/S1473-3099(20)30243-7 doi: 10.1016/S1473-3099(20)30243-7
|








Figures(9) / Tables(1)

Luyu Zhang, Zhaohua Zhang, Sen Pei, Qing Gao, Wei Chen. Quantifying the presymptomatic transmission of COVID-19 in the USA[J]. Mathematical Biosciences and Engineering, 2024, 21(1): 861-883. doi: 10.3934/mbe.2024036







 DownLoad:
DownLoad: